In an earlier essay that covered tunicates, the tunicate larva has two distinction visual action paths, one for phototaxis and one for looming. The two paths use different photoreceptors. Phototaxis photoreceptors are directional with pigment cells blocking light from one direction, while dimming photoreceptors are unidirectional with no shadow from pigment cells.
Looming and dimming are signals of both predators above the animal that block light from the sky, and of obstacles, which also blocks out light from the sky as the animal nears the barrier. In this essay I’ll be focusing on obstacle avoidance using a similar simulation approach as [Zhao et al 2023]. In general, the sky is the brightest, the ground is also light, such as sand, and obstacles are darker. So, if the eye is next to a barrier the average light is dim, while if it’s far from the wall the light is bright because the sky above and the lighter ground below are unobstructed.
Views from the left and right eye for a barrier to the left of an animal. The left is darker because it’s close to the wall, and the right is brighter because it has a clear horizon.
The above screenshot shows a fish-like create with a wall to its left. The left eye is next to a wall, and the right eye views the open field. If the image is reduced to a single average value, the left eye is dimmer, while the right is almost as bright as a full open field. As the first approaches the wall, the image dims rapidly.
Tunicate ascidian dimming
The tunicates (ascidian sea squirts) are the closest non-vertebrate chordates, although evolution has optimized them by removing features, making it difficult to draw direct comparisons to vertebrates [Holland 2016]. The ascidians have a simple larva form that swims for less than 24 hours before settling and becoming a sessile filter feeder.
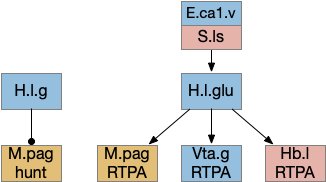
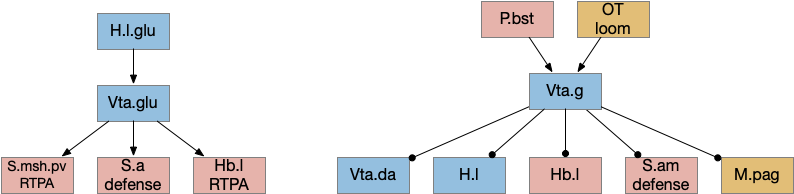
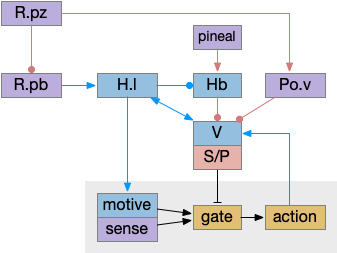
The ascidian larva has a single ocellus (simple, non-image-forming photoreceptor area) has two distinct photoreceptor types and corresponding action paths, one that produces phototaxis and another that responds to rapid dimming [Ryan et al 2016].
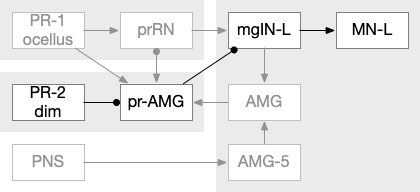
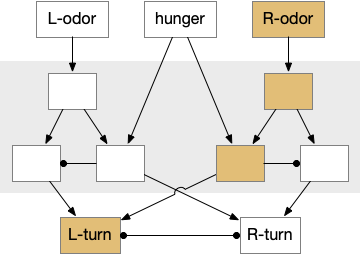
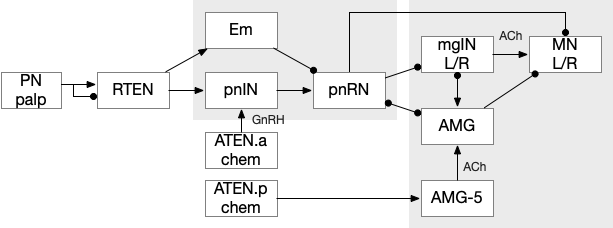
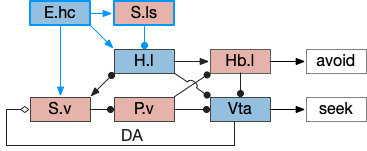
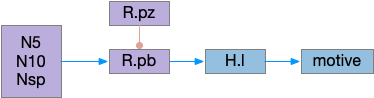
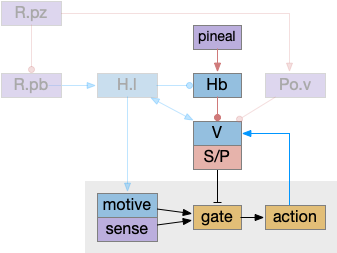
Ascidian nervous system for both phototaxis action path and dimming escape. PR-1 is the directional photoreceptor for phototaxis. PR-2 is the unidirectional photoreceptor for looming escape. mgIN-L and MN-L are motor neurons. AMG is ascending motor feedback.
The above diagram of part of the ascidian larva nervous system, the PR-1 photoreceptors are directional for the top phototaxis path, while PR-2 non-directional photoreceptors produce dimming. The boxes above represent individual neurons, not larger functional groups. MgIN and MN are motor control and motor neurons [Ryan et al 2016].
Larval lamprey primitive eye
Lampreys and hagfish are the only remaining agnathans (non-jawed vertebrates), representing a much larger agnathan vertebrate group that preceded the jawed vertebrates, most of which were filter feeders or sediment feeders [Mallat 2023]. Lamprey larvae are unique among vertebrates in having a primitive non image-forming eye, more like the ascidian ocellus [Bayramov et al 2022]. The adult image-forming retina expands in rings around the more primitive center [Barandela et al 2023].
This central primitive eye is responsive to dimming, and it projects to an equivalent M.pot (pretectum), which handles several optical action paths in zebrafish, including dimming responses, phototaxis, OMR (optomotor reflexes), OKR (optokinetic reflex), and hunting.
Zebrafish retina arborization fields
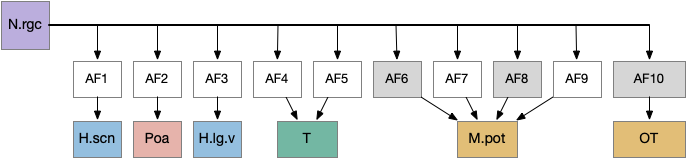
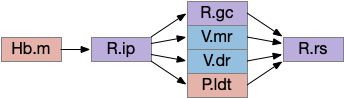
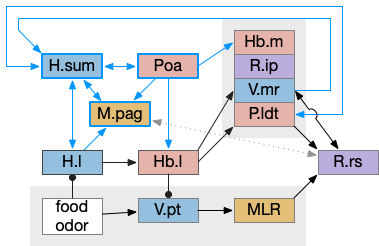
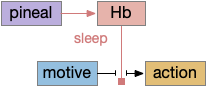
The zebrafish N.rgc (retina ganglion cell) projects to ten distinct AFs (arborization fields), each with a distinct purpose, from AF1 to H.scn (suprachiasmatic nucleus) for circadian timing to AF10 to OT (optic tectum) [Baier and Wullimann 2021]. In most cases distinct N.rgc neuron types project to distinct arborization fields. Even with the largest field AF10 for OT, individual N.rgc neurons project to distinct OT layers. The temporal phototaxis of a previous essay used the projection to AF4 to thalamus to Hb.m (medial habenula, dorsal Hb in zebrafish) [Cheng et al 2017], [Chen and Engert 2014].
The diagram above shows the zebrafish arborization fields and their targets, although the function of many of the targets is not fully known. Dimming fields include AF6, AF8 and OT [Baier and Wullimann 2021], [Temizier et al 2015]. It seems likely that AF6 and AF8 have distinct functionality, although the distinction is not yet known. In lamprey the central ocellus-like photoreceptors project to M.pot, while the outer, lateral image areas project to OT [Cornide-Petronio et al 2011]. The OT dimming response is directional, dimming in one side produces turning [do Carmo et al 2018].
Optic tectum
OT (optic tectum) has the largest arborization and it is the largest nucleus in the midbrain, larger than the entire zebrafish forebrain (cortex / basal ganglia). The optic tectum responds to looming objects and in zebrafish is used for visual hunting [Liu et al 2022]. Since the early vertebrates were filter feeders, the hunting functionality would be unnecessary, leaving obstacle and predator avoidance.
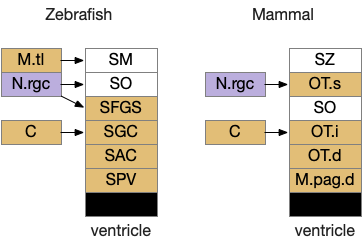
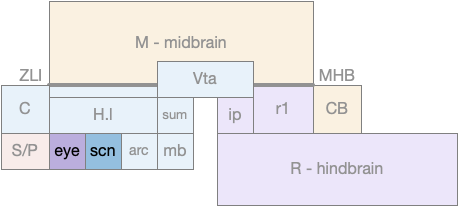
Optic tectum comparison between zebrafish and mammals. C (cortex/pallium), M.pag (periaqueductal grey), M.tl (torus longitudinal), N.rgc (retina ganglion cells), OT (optic tectum)
The optic tectum is layered with retina information arriving in the superficial layers, integrative information from other senses in intermediate layers, and motor actions from the intermediate and deep layers.
The above diagram shows a rough correspondence between zebrafish and mammal optic tectum layers. For simplicity, I’ll use the mammalian names. It’s not clear to me if the PV layer (periventricular layer) of zebrafish is equivalent to M.pag.d (dorsal periaqueductal grey), but I haven’t read any study addressing the physical similarity either as homologous or non-homologous, so it’s probably best to assume the location similarity is merely coincidental.
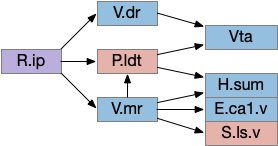
In zebrafish, the OT.s (superficial grey layer, SFGS in zebrafish) itself is layered with each layer receiving distinct N.rgc input [Liu et al 2022]. Dimming input goes to the deepest layer of OT.s [Temizier et al 2015], which is used by OT.d for looming responses [Heap et al 2018]. In mammals, OT.s receives retina input, OT.i produces turn actions, OT.d produces seek and avoid actions, and M.pag.dm produces fast escape from predators.
In vertebrates, OT uses visual expansion (looming) in combination with dimming [Nakagawa and Hongjian 2010], and dimming by itself does not trigger escape [Dunn et al 2016]. However, in the context of the essay’s simulation of a more primitive animal, expansion requires more sophisticated processing from an image-forming eye, which is only available for later vertebrates and not available to even the larval lamprey.
Torus longitudinus
In teleosts (most bony fish), M.tl (torus longitudinal) is a unique nucleus between the left and right OT. M.tl averages the dimming value between the right and left [Folgueira et al 2020]. It also has a sustain role, maintaining behavior after an initial signal.
Torus longitudinus between both OT. M.tl (torus longitudinal), OT (optic tectum).
Interestingly, M.tl is a CB-like (cerebellum-like) structure [Folgueira et al 2020]. Other CB-like ares such as MON (CB-like for LL) and DON (CB-like for electro sensation) act like adaptive filters for the lateral line to cancel out self-motion effects from sensors [Bell et al 2008], [Montgomery et al 2012].
Note that M.pot also communicates with its opposite side through the posterior commissure [Suzuki et al 2015], which could resemble an ancestral visual system. So, although M.tl is directly relevant to the looming response in zebrafish, it may be a specific teleost system, not an indication of an ancestral architecture.
nMLF optical motor output
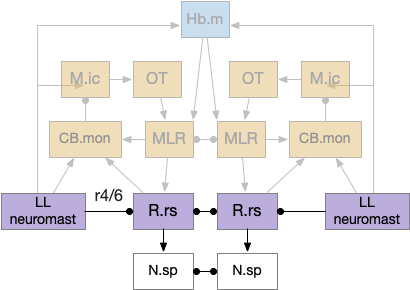
The zebrafish reticulospinal motor control neurons are divided into several groups with distinct action paths. Optical motor output uses M.nmlf (nucleus of the medial longitudinal fasciculus), a midbrain reticulospinal group composed of 20 neurons on each side [Severi et al 2014]. M.nmlf avoidance is distinct from the Mauthner cell startle circuit in r4 in R.mrs. Although the OT looming / dimming can trigger the startle response [Temizer et al 2015], it generally uses the lower-priority M.nmlf [Bhattacharyya et al 2017].
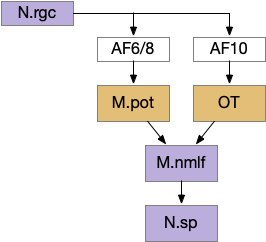
nMLF as the output of the dimming/looming response. AF (retina arborization field), M.pot (pretectum), N.rgc (retina ganglion cell), N.sp (spinal cord), OT (optic tectum).
This direct OT to M.nmlf projection applies to early zebrafish larva. As the fish ages, OT adds projections to R.mrs (middle reticulospinal) in r4-r6 of the hindbrain [Barandela et al 2023], including turning neurons marked by chx10 [Cregg et al 2020]. For this essay, I’m using the simpler early projection to M.nmlf.
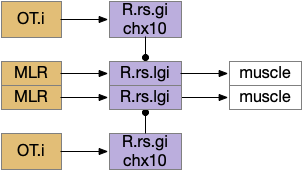
Looming can produce zebrafish O-bends (u-turns) as well as directional turns [Portugues and Engert 2009], [Marques et al 2018]. For this essay, I’m assuming that M.pot produces a base O-bend command that the OT can modify by choosing a turn direction. This split between motivation and turning also occurs in R.mrs, where MLR (midbrain locomotive region) produces a non-directional forward movement, while chx10 neurons in R.mrs receive OT turning commands for looming [do Carmo et al 2018], [Cregg et al 2020].
Simulation
This essay’s simulation uses dimming as an obstacle avoidance system, similar to the simulation in [Zhao et al 2023], but with a minimal dimming input. The essay’s simulation condenses the input to the simplest dimming structure, where each eye has only a single averaged luminance value. The retina also calculates a dimming value as the difference between the current luminance and the previous value. Although the vertebrate retina uses distinct unsigned ON and OFF channels, the simulation uses a single signed value.
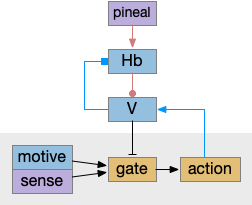
The looming module triggers a looming response when the dimming value passes a threshold as a proportion of the current luminance. This part of the model represents M.pot (pretectum). If no further information is available, the looming triggers a u-turn (O-bend in zebrafish) using M.nmlf.
If the left and right eyes have a difference in brightness, the model converts the u-turn into a left turn or right turn. This part of the model represents the OT’s dimming response. Like the M.pot output, this OT turn signal uses M.nmlf, as in the early zebrafish larva.
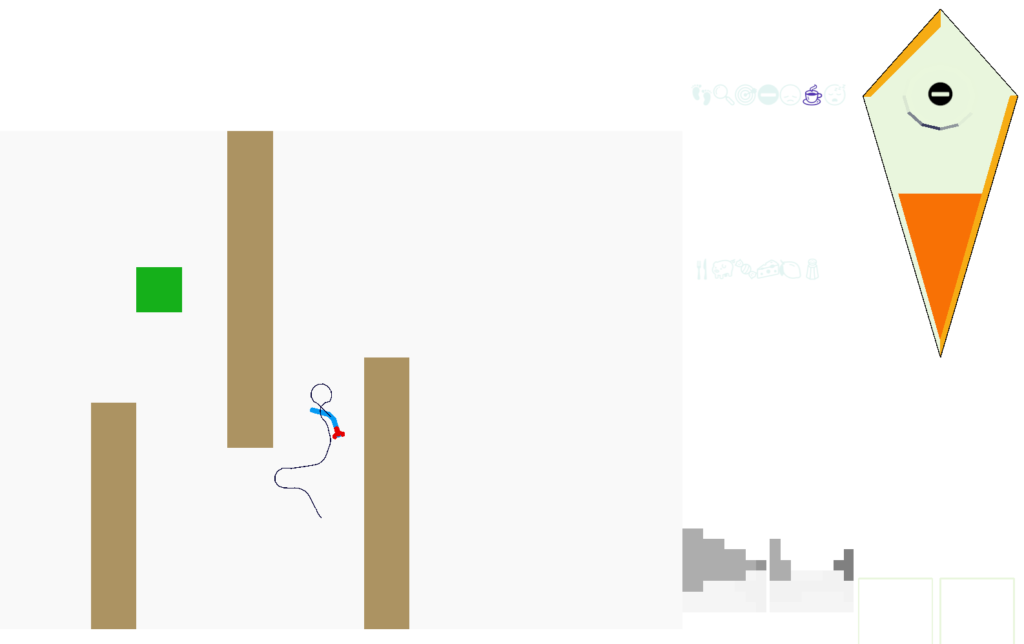
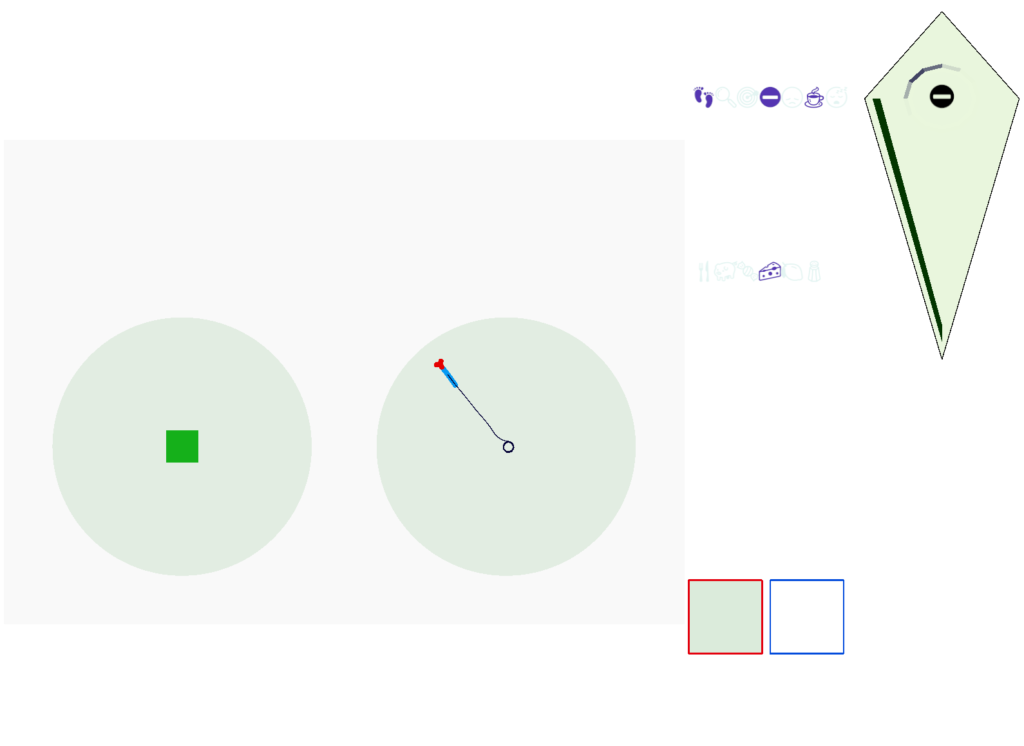
Screenshot showing the animal avoiding a wall to its left. The left and right retina displays are for human viewing.
The above screenshot shows the animal avoiding an obstacle to its left. The two low-resolution images at the lower right are for human viewing and are higher resolution than the animal uses. The animal itself only uses a single averaged value for each eye. This view from the left eye is dominated by the wall, which blocks the light. The right eye mostly sees a clear view to the horizon.
Discussion
Qualitatively, the system works surprisingly well despite its simplicity. In some of the narrow corridors the u-turn behavior will reverse out of the corridor, and the entrance to the corridors is something of a barrier because only the center of the corridor will avoid triggering avoidance.
The model doesn’t adjust speed, which is an interesting potential improvement. If the animal slowed near obstacles, raised the threshold for obstacle avoidance, and reduced the turn angles, it might more easily navigate corridors. Since searching already has a roam vs dwell mode for ARS (area restricted search), triggered by serotonin, a slow-moving obstacle avoidance mode could use the same mechanism. V.dr (dorsal raphe serotonin) does reduce looming defense [Huang et al 2017]. Alternatively, since OT.d looming does habituate [Lee et al 2020], that habituation could reduce the excessive u-turning of the model. H.lgn.v (ventral lateral geniculate nucleus), which responds to overall light levels, can also inhibit the looming response [Fratzl et al 2021].
When seeking an odor, vertebrate swimming undulates left and right, naturally moving the nose perpendicular to the body motion. This lateral motion can help navigation if odor sampling can be coordinated with the movement, enabling a spatiotemporal gradient calculation along the path of the nose movement. This lateral sampling over time is called klinotaxis (“leaning navigation”) or weathervaning.
Essay 24 and essay 25 explored head-direction navigation as inspired by the fruit fly Drosophila fan-shaped body and ellipsoid body. The idea was to use head direction to translate egocentric movement into an allocentric memory of past samples, independent of the current body direction. In contrast, klinotaxis uses an egocentric system, where the lateral motion is relative to the current direction, not an independent, compass or map-like system.
Klinotaxis in Drosophila larva and C. elegans
Klinotaxis has been largely studied in the fruit fly Drosophila larva and the roundworm C. elegans. Drosophila larva have a distinct “cast” movement, where they pause and wave their heads side to side, either a single time (1-cast) or multiple times (n-cast) [Zhao et al 2017]. Larva movements break down into five major types [Gomez-Marin and Louis 2014]:
Forward
Backward
Stop
Turn
Cast
C. elegans has two major seek movements: pirouettes and weathervaning [Lockery 2011]. Pirouettes are a u-turn when the animal is moving away from the odor. Weathervaning is a side-to-side head movement that manages turning.
Both systems are temporal gradient systems, requiring measurements at different times and a memory of the older measurement [Chen X and Engert 2014]. Klinotaxis requires a basic form of memory [Karpenko et al 2020], but the comparison can be a simple ON or OFF result [Lockery 2011]. Pirouetts use a gradient parallel to body motion and reverse direction when the animal is moving away from the odor [Iino and Yoshida 2009]. Weathervaning uses a gradient perpendicular to body motion, measured with a lateral head movement [Lockery 2011].
This klinotaxis contrasts with a bilateral spatial navigation that compares two lateral sensors [Chen X and Engert 2014], such as bilateral eyes, ears, or nostrils. In Drosophila larva, odor turning is proportional to the lateral gradient more than the parallel gradient [Martinez 2014]. The odor navigation is not simply bilateral because disabling one side of O.sn (olfactory sensory neuron) only minimally impairs navigation [Gomez-Marin and Louis 2014].
As a slight digression, let’s return to the adult Drosophila navigation, because the structure can be a useful analogy for understanding vertebrate klinotaxis navigation, despite using a different allocentric system.
Adult Drosophila FSB
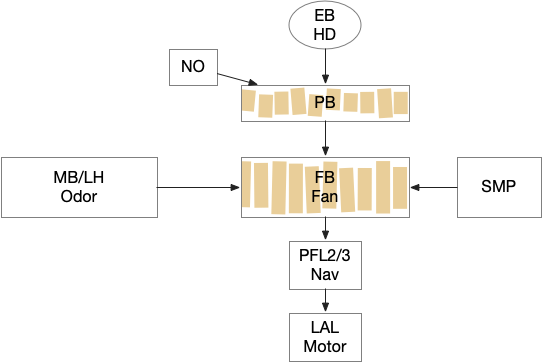
Below is a rough sketch of the Drosophila navigation circuit, focused on the fan-shaped body [Hulse et al 2021]. The ellipsoid body (EB) and protocerebral bridge (PB) calculate head direction and sort it into 18 columns. This head direction is allocentric, independent of the animal’s current direction, like a compass direction or a map. Input from odor areas like the mushroom body (MB) and lateral horn (LN) are organized into 9 rows. The fan-shaped body combines these 18 head direction columns and 9 sense data rows into a memory table.
Drosophila navigation, focusing on head direction from PB, odor data from MB and LH, and allocentric table of FB. EB ellipsoid body, FB fan-shaped body, LH lateral horn, MB mushroom body.
Motor navigation reads out from the fan-shaped-body table. These motor commands include left and right, but also include a separate u-turn command [Westeinde et al. 2022]. Although this allocentric navigation system differs from egocentric klinotaxis, its motor output includes both the left vs right from weathervaning and the u-turn from pirouette.
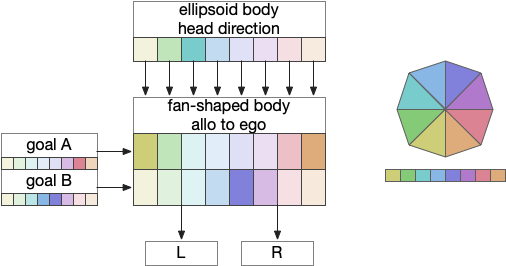
The previous essay 24 and essay 25 attempts followed this model. As the animal moves in space, the model saved the forward odor gradient according to the current head direction. By comparing stored values for other head directions, the animal would improve its heading toward the direction with the strongest odor.
The fan-shaped body then becomes a record of samples of all the older directions that the animal had measured. Output is then calculated for left (PFL3L), right (PFL3R), and u-turn (PFL2) signals. [Westeinde et al 2024]. The current head direction is represented as a sinusoidal neural pattern and combined with the stored values to produce an output.
This system was only partially successful for the essay. Although it was an improvement over no memory, because the animal was continually moving in space, the table was always obsolete. Even when the table memory times out to represent loss in accuracy as the animal moves, the rapid obsolescence made navigation difficult, particularly as the animal neared the target.
So, this essay simplifies the circuit and lowers the ambition. Instead of trying to record every direction and keeping perfect allocentric compass direction, the animal could simple save its left and right oscillation as it swims naturally.
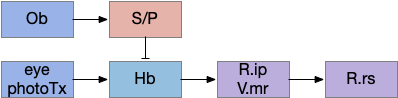
Vertebrate Hb.m and R.ip
The vertebrate Hb.m (medial habenula) to R.ip (interpeduncular nucleus) is used for phototaxis [Chen X and Engert 2014], Chemotaxis [Chen WY et al 2019] and thermotaxis [Palieri et al 2024]. In a clever experiment creating a virtual light circle, Chen and Engert shows that the zebrafish phototaxis is not simply comparing light between the eyes for a spatial gradient (tropotaxis) but is a temporally-based gradient (klinotaxis), relying on a short term memory of the previous light. This phototaxis uses the Hb.m to R.ip circuit [Chen X and Engert 2014].
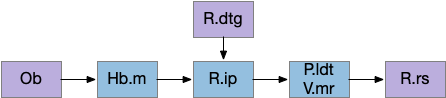
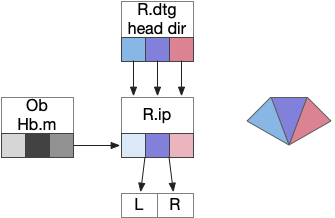
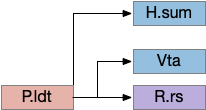
Head direction from R.dgt (dorsal tegmental nucleus) tiles R.ip vertically [Petrucco et al 2023], while olfactory and light input is organized horizontally [Chen WY et al 2019], [Zaupa et al 2021]. After combining the odor with the head direction and comparing with the stored values, it sends motor commands to R.rs (reticulospinal) using P.ldt (laterodorsal tegmental nucleus) and V.mr (median raphe). The vertebrate R.ip has 6 columns of head direction input from R.dtg, resembling the Drosophila fan-shaped body, but instead of 18 columns for the fan-shaped body, R.ip only has 6, three to a side [Petrucco et al 2023].
Essay 25 explored a model which used the Drosophila fan-shaped body allocentric navigation in R.ip with some limited but not overwhelming success. Instead, this essay will try a different interpretation, where R.ip is only storing side to side weathervaning of the head while swimming, instead of a full 360 degree table like Drosophila.
Vertebrate klinotaxis
As a different approach, suppose the head direction to R.ip is not an allocentric map-making coordinator as in the adult Drosophila, but a simpler egocentric weathervaning or casting coordinator, storing only the lateral gradient from head direction changes from natural swimming, or possibly deliberate larger turns like casting to gather wider lateral gradient information.
Klinotaxis simplifies the need for precise head direction. Instead of the Drosophila 18 head direction columns calibrated to the outside world, we use only three, two lateral and one central, that only require motor efference copies of left and right muscle turns. Studies from the zebrafish R.ip suggest three columns to a side, which isn’t connected to the vestibular system [Petrucco et al 2023]. To me, this suggests to me that the head direction might not be an allocentric signal that requires precise direction, but a simple egocentric lateral measurement, which doesn’t need vestibular information.
The above diagram illustrates the system. Olfactory samples arrive through Hb.mand head direction arrives from R.dtg. Like the Drosophila fan-shaped body, R.ip combines odor samples with lateral head movement into a simple memory table, and it reads out left and right motor commands. A similar system can save odor measurements parallel to body movement, using velocity instead of head direction, to trigger a u-turn when the animal is moving away from the odor.
Discussion
Compared to the parallel-only gradient, allocentric system of essay 25, this lateral navigation is far simpler and more effective. Even with only three bins compared to the 8 bins in essay 25, the lateral weathervaning turned out to be more effective and less brittle. If R.ip does implement a lateral klinotaxis system like this essay, it’s plausible that the 6 directions reported by [Westeinde et al 2024] are sufficient for accurate seek navigation. In contract, those 6 directions seem insufficient for an allocentric navigation compared to the Drosophila 18 directions.
Interestingly, the pirouette also highly effective, even without lateral klinotaxis. In the simulation, when the animal moved away from the odor source, it makes a u-turn. This system served to ratchet the animal closer and closer to the target. Even when most of the movement was random, the pirouette locks in any improvement. Pirouette itself is also simple, only requiring two averages: a short average and a long average, where a short average tracks the odor across a single swim cycle and a long average uses two swim cycles. When the short average has a stronger odor value than the long average, the animal is moving toward the odor.
In both cases, the simulation used a binary OFF for the motor command instead of attempting finer precision from the gradient. This simple OFF strategy was sufficient for the simulation. A C. elegans study suggested that ON-OFF coding was energy efficient, and the worm rarely orients perfectly to the gradient [Lockery 2011].
The ascidian circuit in essay 30 had an interesting dopamine subcircuit that looks like an indirect search, where the ascidian coronet cells modulate the underlying phototaxis and geotaxis circuits. While the function of the coronet cells is unknown, if these cells are another seeking system like following an odor, then the coronet sub circuit follows odor by modulating different seek circuits: phototaxis and geotaxis.
Ascidian analogy
Tunicates are the closest non-vertebrate chordates evolutionarily, but they have developed in vastly different directions from the vertebrates, and likely very differently from the shared common ancestor [Holland 2015]. The ascidian tunicates, which are the most studied tunicates, live their asul life as sessile filter feeders like sponges. Their eggs hatch in only 20 hours and their brief tadpole form lasts only for a few hours, just enough to swim and disperse to find a likely permanent settlement place. Their locomotive strategy is to swim up using geotaxis in the morning and swim down using phototaxis in the afternoon. If they’re lucky enough to find a ledge, they swim up into the ledge’s shadow to settle because hanging like a bat from a ledge offers more protection from some predators than resting on the ocean floor [Zega et al 2006].
As would be expected from a 20-hour brain, the navigation circuit is fairly simple. There are two distinct action paths, one for geotaxis using a heavy pigment cell and one for phototaxis using photoreceptors and another pigment cell as a shadow to provide photo-directionality. The two action paths are connected, where dimming produces upward swimming [Bostwick et al 2020].
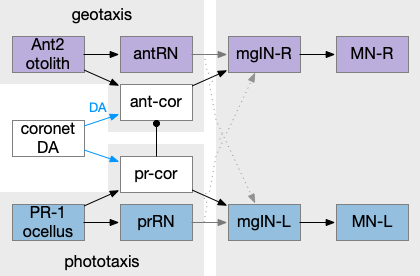
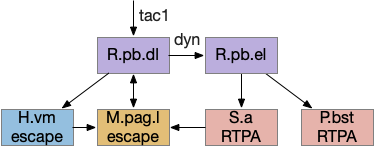
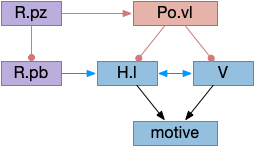
Ascidian tadpole sub circuit for geotaxis and phototaxis. The horizontal neurons are the main action paths. The coronet DA cells modulate the action paths.
In the above diagram, the geotaxis action path starts from the otolith (“ear stone”) receptor ant2, which is functionally similar to the vestibular system (but not related), passes input to antenna relay neurons (antRN) and then to the right side motor neurons (mgIN-R and MN-r) [Ryan et al 2016]. Similarly, the phototaxis action path starts from the ocellus (eyespot) to the phototaxis relay (prRN) and to the left motor neurons, providing an opposing direction from geotaxis. Importantly for the following discussion, each path has a weak connection to the opposite direction, possibly to add some stochasticity to the movement to improve dispersion of the many tadpoles.
The function of the coronet cells is unknown, although they have some genetic connection the palp sensory cells [Cao et al 2019]. Other papers compare the corona cells to dopamine cells in the hypothalamus and Ob (olfactory bulb) [Horie et al 2018] or ancestral photo-hypothalamus and retina [Sharma et al 2019], possibly related to the fish saccus vasculosus area of the hypothalamus, responsible for some circadian behavior. However, the ascidian tadpole has lost circadian clock genes, which argues against circadian timing [Chung et al 2023]. The coronet cells can accumulate serotonin and the DA might promote onset of metamorphosis [Razy-Kraika et al 2012]. So, the coronet may be involved in triggering metamorphic changes at twilight, which causes the tadpole to dive to deeper waters [Lemaire et al 2021].
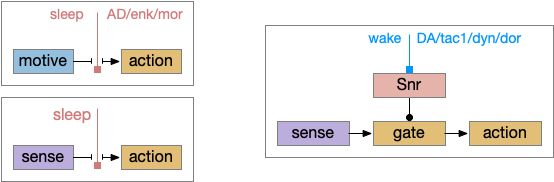
Whatever the source, the interesting thing about the circuit is that it’s an indirect modulation of underlying taxis action paths. The action of the coronet is gating or modulatory. While this coronet circuit is not homologous to the basal ganglia, using it as an analogy may be useful. For example, dopamine is a sleep / wake signal for the basal ganglia [Vetrivelan et al 2010]. Because low dopamine reduces basal ganglia activity both at the striatum input layer and the Snr (substantia nigra pars reticulata) output layer, it’s an effective sleep controller.
Indirect chemotaxis
Consider indirect chemotaxis, where the animal is seeking toward the odor, but the underlying action path is phototaxis or geotaxis, like the ascidian circuit above. If the animal detects an odor, it increases the current direction. In other words, the current direction is toward or near a food odor. This strategy is like the e. coli tumble-and-run strategy, where the bacteria runs further when the odor gradient is increasing.
Consider the basal ganglia as an analogy. For example, Ob has some dopamine interneurons (Ob.sac – short axis cells) that project to S.ot (olfactory tubercle) [Burton 2017], a portion of the stratum focused on olfactory input. For the corollary of the phototaxis path, consider the Hb.m (medial habenula) phototaxis path [Zhang et al 2017].
Hypothetical indirect seek circuit where chemotaxis uses an underlying phototaxis to hunt for food. Hb (habenula), Ob (olfactory bulb), P (pallidum), R.ip (interpeduncular nucleus), R.rs (reticulospinal motor neurons), S (striatum), V.mr (median raphe).
When the odor is detected, Ob enables the basal ganglia, which enhances the phototaxis path. If the odor isn’t detected, the default semi-suppressed behavior means the direction is semi-random. This indirect control would allow for seeking odor when the underlying navigation is phototaxis and geotaxis.
Discussion
After writing this description. I think this model may be a bit sketch for something like chemotaxis, although it’s a reasonable model for sleep. Because I’m not sure the idea is likely to be productive, I’m holding off on doing any implementation, but writing down the description in case it makes sense later.
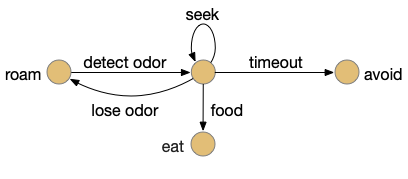
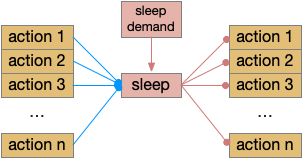
Let’s return to the task of essay 16 on give-up time in foraging, which covered food search with a timeout. At first the animal uses a general roaming search and if it smells a food odor, it switches to a targeted seek following the odor with chemotaxis. If the animal finds food in the odor plume, it eats the food, but if it doesn’t find food, it will eventually give up and avoid the local area before returning to the roaming search.
Search state machine. Roam is the starting state, switching to seek when it detects odor, and switching to avoid after a timeout.
For another attempt at the problem, let’s take the striatum (basal ganglia) as implementing the timeout portion of this task using the neurotransmitter adenosine as a timeout signal and incorporating the multiple action path discussion from essay 30 on RTPA. Adenosine is a byproduct of ATP breakdown and is a measure of cellular activity. With sufficiently high adenosine, the striatum switches from the active seek path to an avoidance path. These circuits are where caffeine works to suppress the adenosine timeout, allowing for longer concentration.
Mollusk navigation
As mentioned in essay 30, the mollusk sea slug has a food search circuit with a similar logic to what we need here. The animal seeks food odors when it’s hungry, but it avoids food odors when it’s not hungry [Gillette and Brown 2015].
Mollusk food search circuit, illustrating a hunger-modulated switchboard. When the animal is not hungry, the switchboard reverses the odor to motor links turning it away from food.
This essay uses the same idea but replaces the hunger modulation with a timeout. When the timeout occurs, the circuit switches from a food seek action path to a food avoid action path.
Odor action paths
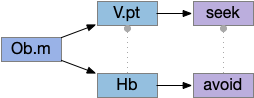
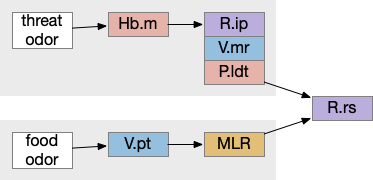
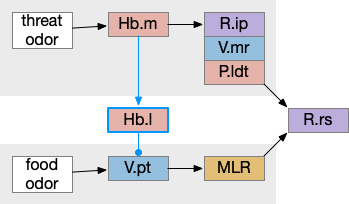
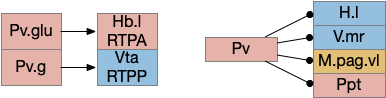
Two odor-following actions paths exist in the lamprey, one using Hb.m (medial habenula) and one using V.pt (posterior tuberculum). The Hb.m path is a chemotaxis path following a temporal gradient. The V.pt path projects to MLR (midbrain locomotor region), but The lamprey Ob.m (medial olfactory bulb) projects to both Hb.m (medial habenula) and to V.pt (posterior tuberculum), which each project to different locomotor paths [Derjean et all 2010], Hb.m to R.ip (interpeduncular nucleus) and V.pt to MLR (midbrain locomotor region). The zebrafish also has Ob projections to Hb and V.pt [Imamura et al 2020], [Kermen et al 2013].
Dual odor-seeking action paths in the lamprey and zebrafish. Hb (habenula), Ob.m (medial olfactory bulb), V.pt (posterior tectum).
Further complicating the paths, the Hb.m itself contains both an odor seeking path and an odor avoiding path [Beretta et al 2012], [Chen et al 2019]. Similarly Hb.m has dual action paths for social winning and losing [Okamoto et al 2021]. So, this essay could use the dual paths in Ob.m instead of contrasting Ob.m with V.pt, but the larger contract should make the simulation easier to follow.
This essay’s simulation makes some important simplifications. The Hb to R.ip path is a temporal gradient path used for chemotaxis, phototaxis and thermotaxis. In a real-world marine environment, odor diffusion and water turbulence is much more complicated, producing more clumps and making a simple gradient ascent more difficult [Hengenius et al 2012]. Because this essay is only focused on the switchboard effect, this simplification should be fine.
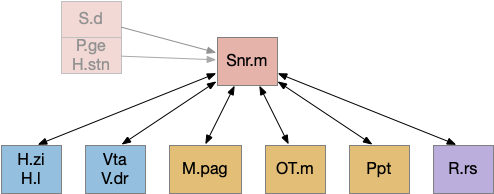
Striatum action paths with adenosine timeout
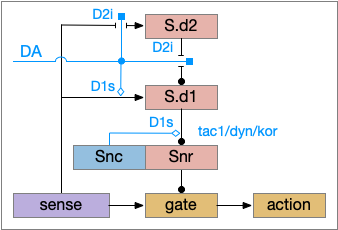
The timeout circuit uses the striatum, which has two paths: one selecting the main action, and the second either stopping the action, or selecting an opposing action [Zhai et al 2023]. The two paths are distinguished by their responsiveness to dopamine with S.d1 (striatal projection with D1 G-s stimulating) or S.d2 (striatal projection with D2 G-i inhibiting) marking the active and alternate paths respectively. This model is a simplification of the mammalian striatum where the two paths interact in a more complicated fashion [Cui et al 2013].
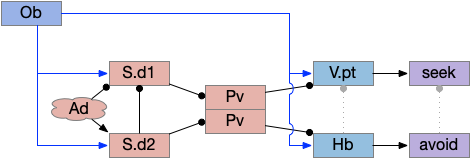
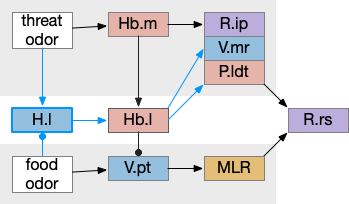
Essay odor seek with timeout circuit. The seek path flows from Ob, through S.d1 to P.v to V.pt. The avoid path flows from Obj, though S.d2 to Pv. to Hb. Ad (adenosine), Hb (habenula), Ob (olfactory bulb), Pv (ventral pallidum), S.d1 (striatum D1 projection neuron), S.d2 (striatum D2 projection neuron), V.pt (posterior tuberculum)
As mentioned, the two actions paths are the seek path from Ob to V.pt and the avoid path from Ob to Hb. For the timeout and switchboard, the Ob has a secondary projection to the striatum. Although this circuit is meant as a proto-vertebrate simplification, Ob does project to S.ot (olfactory tubercle) and to the equivalent in zebrafish [Kermen et al 2013].
The timeout is managed by adenosine, which is a neurotransmitter derived from ATP and a measure of neural activity. The striatum has three sub-circuits for this kind of functionality, which I’ll cover in order of complexity.
S.d1 and adenosine inhibition
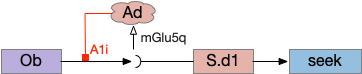
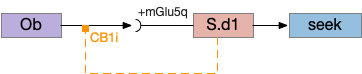
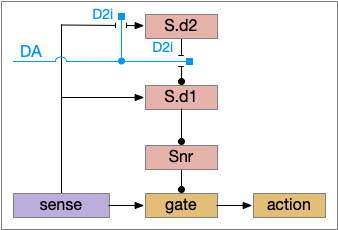
The first circuit only uses the direct S.d1 path and adenosine as a timeout mechanism. When the animal follows an odor, the Ob to S.d1 signal enables the seek action. As a timeout, ATP from neural activity degrades to adenosine and the buildup of adenosine is a decent measure of activity over time. The longer the animal seeks, the more adenosine builds up. Of the Ob projection axis contains an A1i (adenosine G-i inhibitory) receptor, the adenosine will inhibit the release of glutamate from Ob, which will eventually self-disable the seek action.
S.d1 action path inhibited by adenosine buildup as a timeout. A1i (adenosine G-i inhibitory receptor), Ad (adenosine), mGlu5q (metabotropic glutamate G-q receptor), Ob (olfactory bulb), S.d1 (D1-type striatal projection neuron)
In practice, the striatum uses astrocytes to manage the glutamate release. An astrocyte that envelops the synapse measures glutamate release with an mGlu5q (metabotropic glutamate with G-q/11 binding) receptor and accumulates internal calcium [Cavaccini et al 2020]. The astrocyte’s calcium triggers an adenosine release as a gliotransmitter, making the adenosine level a timeout measure of glutamate activity. The presynaptic A1i receptor then inhibits the Ob signal. The timeframe is on the order of 5 to 20 minutes with a recovery of about 60 minutes, although the precise timing is probably variable. Interestingly, the time-out is a log function instead of linear measure of activity [Ma et al 2022].
This circuit doesn’t depend on the postsynaptic S.d1 firing [Cavaccini et al 2020], which contrasts with the next LTD (long term depression) circuit which only inhibits the axon if the S.d1 projection neuron fires.
S.d1 presynaptic LTD using eCB
S.d1 self-activating LTD uses retrotransmission to inhibit its own input using eCB (endocannabiniods) as a neurotransmitter. Like the astrocyte in the previous circuit, S.d1 uses a mGlu5q receptor to trigger eCB release, but also require that S.d1 fire, as triggered by NMDA glutamate receptor. The axon receives the eCB retrotransmission with a CB1i (cannabinoid G-i inhibitory) receptor and trigger presynaptic LTD [Shen et al 2008], [Wu et al 2015]. Like the previous circuit, the timeframe seems to be on the order of 10 minutes, lasting for 30 to 60 minutes.
S.d1 LTD circuit. A coincidence of glutamate detection with mGlu5q and S.d1 activation with NMDA triggers eCB release, which activates CB1i leading to presynaptic LTD. CB1i (cannabinoid G-i inhibitory receptor), mGlu5q (glutamate G-q receptor), Ob (olfactory bulb), S.d1 (striatum D1-type projection neuron).
This circuit inhibits itself over time without using adenosine or astrocytes. In the full striatum circuit, high dopamine levels suppress this LTD suppression, meaning that dopamine inhibits the timeout [Shen et al 2008].
The next circuit adds the S.d2 path, which uses adenosine and self-activity to trigger postsynaptic LTD.
S.d2 postsynaptic LTP via A2a.s
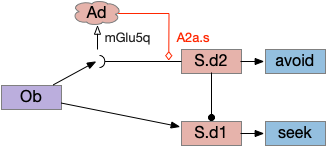
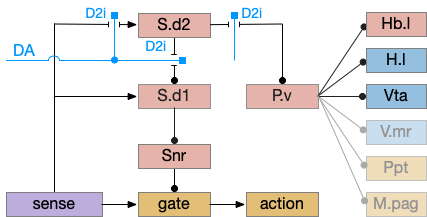
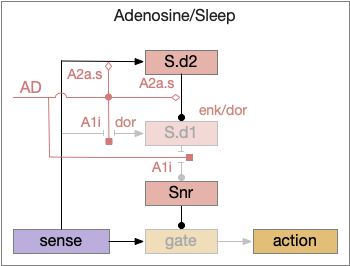
Consider a third circuit that has the benefits of both previous circuits because it uses adenosine as a timer managed by astrocytes and is also specific to postsynaptic activity. In addition, it allows for a second action path, changing the circuit from a Go/NoGo system to a Go/Avoid action pair. This circuit uses LTP (long term potentiation) on the S.d2 striatum neurons.
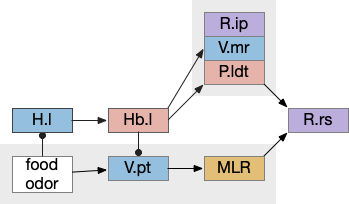
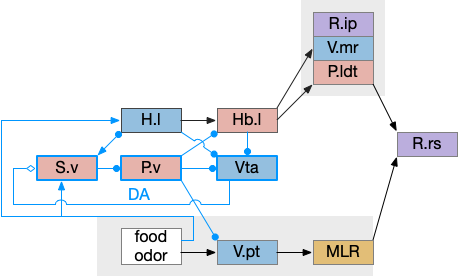
Timeout circuit using postsynaptic LTD at the S.d2 neuron and adenosine as a timeout signal. As adenosine accumulates, it stimulates S.d2, which both disables S.d1 and drives the avoid path. A2a.s (adenosine G-s stimulatory receptor), Ad (adenosine), mGlu5q (glutamate G-q metabotropic receptor), Ob (olfactory bulb), S.d1 (striatum D1-type projection neuron), S.d2 (striatum D2-type projection neuron)
When the odor first arrives, Ob activates the S.d1 path, seeking toward the odor. S.d1 is activated instead of S.d2 because of dopamine. In this simple model, the Ob itself could provide the initial dopamine like c. elegans odor-detecting neurons or the tunicate’s coronal cells or the dual glutamate and dopamine neurons in Vta (ventral tegmental area).
As time goes on, adenosine from the astrocyte builds up, which activates the S.d2 A2s.a (adenosine G-s stimulatory receptor) until it overcomes dopamine suppression and increases the S.d2 activity with LTP [Shen et al 2008]. Once S.d2 activates, it suppresses S.d1 [Chen et al 2023] and drives the avoid path.
The combination of these circuits looks like it’s precisely what the essay needs.
Simulation
In the simulation, when the animal is hunting food and finds a food odor plume, it directly seeks toward the center and eats if it find food. In the screenshot below, the animal is eating.
Simulation showing the animal eating food after seeking the odor plume.
Satiation disables the food seek. This might sound obvious, but hunger gating of food seeking requires specific satiety circuits to any seek path that’s food specific, which means the involvement of H.l (lateral hypothalamus) and related areas like H.arc (arcuate hypothalamus) and H.pv (periventricular hypothalamus). And, of course, the simulation requires simulation code to only enable food odor seek when the animal is searching for food.
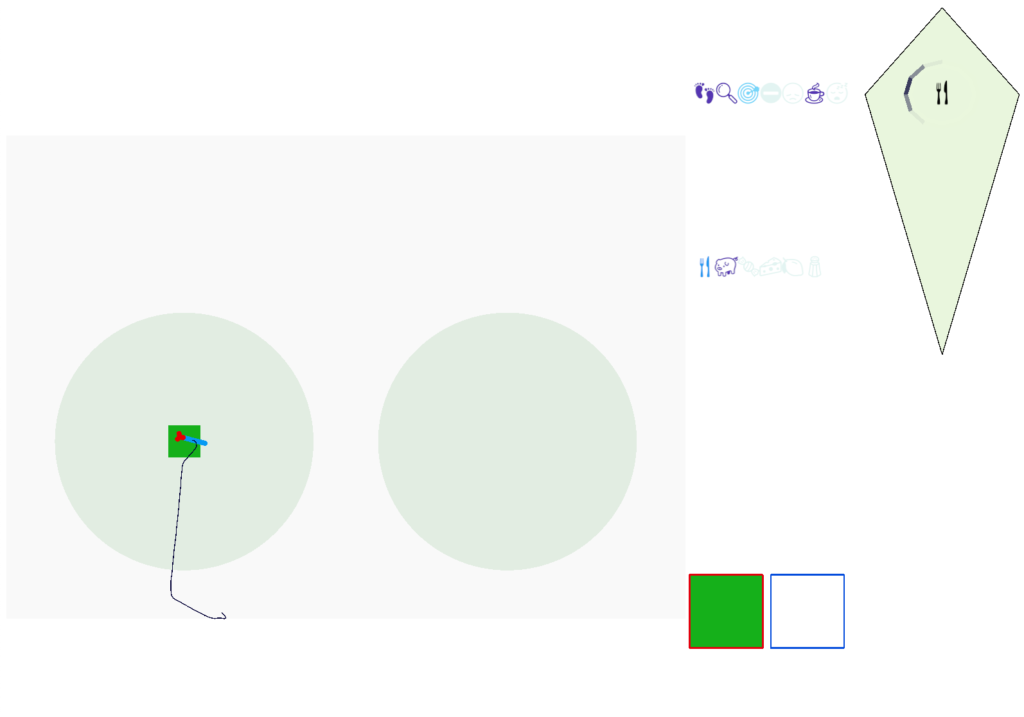
The next screenshot shows the central problem of the essay, when the animal seeks a food odor but there’s no food at the center.
Screenshot showing the animal stuck in the middle of the food odor plume before the timeout.
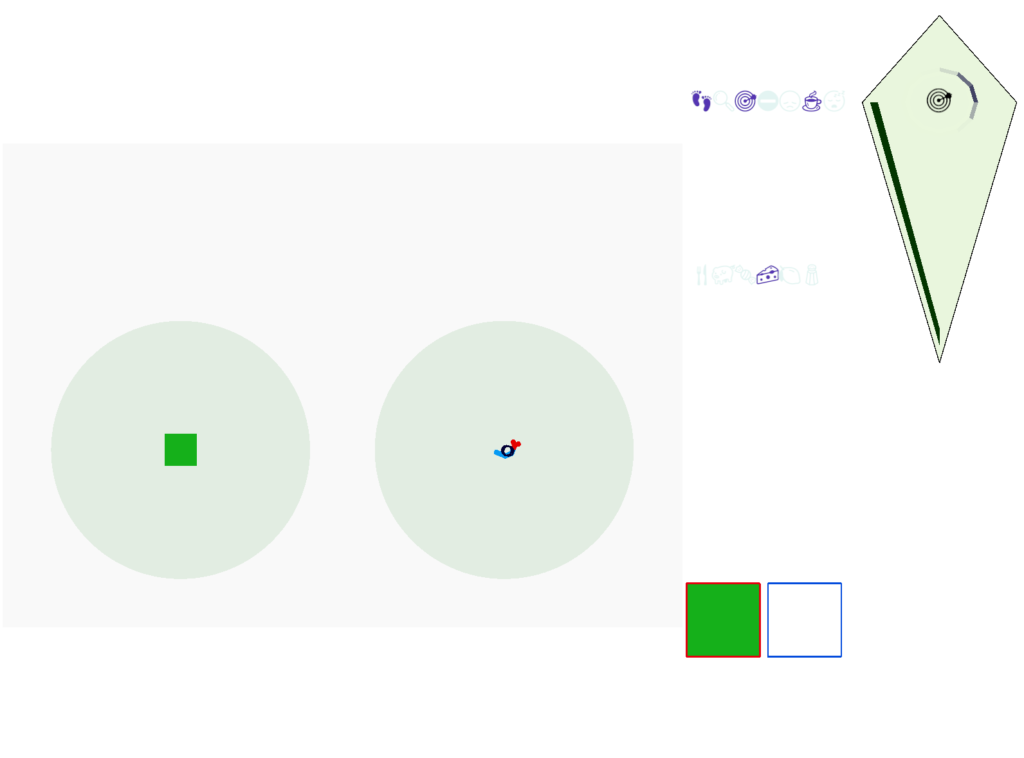
Without a timeout, the animal circles the center of the food odor plume endlessly. After a timeout, the animal actively leaves the plume and avoid that specific odor until the timeout decays.
Screenshot showing the animal escaping from the odor plume after the timeout.
This system is somewhat complex because of the need for hysteresis. A too-simple solution with a single threshold can oscillate, because as soon as the animal starts leaving the timeout decays, which then re-enables the food-seek, which then quickly times out, repeating. Instead, the system needs to make re-enabling of the food seek more difficult after a timeout.
But that adds a secondary issue because if food seek is a lower threshold, then the sustain of seek needs to raise the threshold while the seek occurs. So, the sustain of seek needs a lower threshold than starting seek. This hysteresis and seek sustain presumably needs to be handled by the actual striatum circuit.
Discussion
I think this essay shows that using the stratum for an action timeout for food seek is a plausible application. The circuit is relatively simple and is effective, improving search by avoiding failed areas.
However, the simulation does raise some issues, particularly hysteresis problem. If the striatum does provide a timeout along these lines, it must somehow solve the hysteresis problem. While the animal is seeking, the ongoing LTP/LTD inhibition should use a high threshold to stop seeking, but once avoidance starts, there needs to be a high threshold to return to seeking to avoid oscillations between the two action paths.
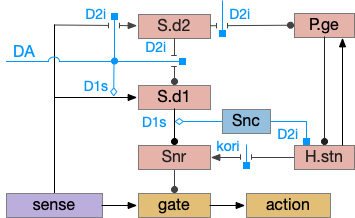
Because LTD/LTP is a relatively long chemical process (minutes) internal to the neurons, as opposed to an instant switch in the simulation, the delay itself might be sufficient to solve the oscillation problem. It’s also possible that some of the more complicated parts of the circuit, such as P.ge (globus pallidus) and its feedback to the striatum or H.stn (subthalamic nucleus) might affect the sustain of seek or breaking it and so control the hysteresis problem.
The simulation also reinforced the absolute requirement that action paths need to be modulated by internal state like hunger. For the seek paths, both Hb.m and V.pt are heavily modulated by H.l and other hypothalamic hunger and satiety signals.
As expected, the simulation also illustrated the need for context information separate from the target odor. While the food odor is timed out, the animal can’t search the other odor plume because this essay’s animal can’t distinguish between the odor plumes, and therefore avoids both odors. With a long timeout and many odor plumes, this delays the food search. A future enhancement is to add context to the timeout. If the animal can timeout a specific odor plume, it can search alternatives even if the food odor itself is identical.
I’m looking to improve the foraging algorithm with an idea from essay 17, which suggested that when the foraging fails, the animal should avoid the failed area. The foraging task uses an odor cue to seek food. Currently when the model gives up (times out), it disables seeking, but doesn’t actively avoid the current place, but returns to the wide-ranging roaming search.
For now, I’m still avoiding memory, but consider the alternating T-maze used in rodent behavior [Deacon and Rawlins 2006]. Mice are released at the base of the T and choose one of the directions to search for food. If the experiment repeats (by picking up the mice and restarting) mice will tend to explore the unexplored end first.
But for our foraging task, let’s use the same device for a different purpose. Instead of repeating the experiment by unnatural teleportation, consider the simpler problem of foraging with this device as an environment.
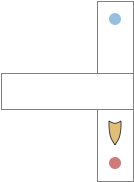
T-maze exploration. Food might be at either the red dot or the blue dot.
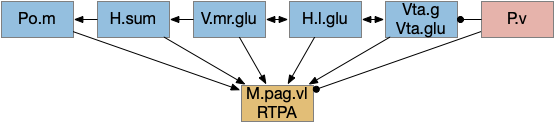
When rodents are foraging and reach one end, they will reverse and search the other end. Because rodents are far more advanced than the toy model, they can remember which arm of the maze they’ve already explored. But consider a simpler sub-strategy that uses RTPA (real-time place avoidance) where the animal temporarily avoids the current area or areas associated with food. By actively avoiding the already-explored area, the animal will save time by avoiding repeated searching.
A difficulty in finding the neural correlates of RTPA is the great diversity of reasons for RTPA, and circuits even in the brainstem. There are many reasons for place avoidance:
Startle: reflex escape
Escape from an imminent predator
Escape from an environment hazard (CO2, temperature)
Avoiding innate cues (predator odors)
Avoiding learned cues (CPA conditioned place avoidance)
Search optimization: avoiding already searched areas
Because this topic is large and the number of circuits is also large, I’ll start with a more abstract view to provide some context for a later dive into details. The two architectures will be a set of labeled path seek and avoidance circuits, and a secondary consensus circuit to coordinate the labeled paths.
Labeled path
A labeled path architecture uses individual circuit paths for each behavior and sense, as opposed to bringing all stimuli into a central node with a general decision algorithm [Helmbrecht 2018]. (Helmbrecht uses “labeled line,” which conflicts with the fish “lateral line” sense.) As least to some extend, the brainstem is designed around labeled paths, which is particularly evident if using the chimera model of the bilateral brain [Tosches and Arendt 2017].
The chimera model posits that brains of bilateral animals combine features from apical (unilateral) and bilateral (“blastoporal” in their terminology because they focus on zooplankton larvae). The apical mode is associated with the front of the brain, such as the hypothalamus, and its locomotion is temporally gradient based, like the tumble-and-run of bacteria. The bilateral mode is more reflexive, turning left if touched on the right, like Braitenberg vehicles [Braitenberg 1984]. Apical systems include olfactory search and phototaxis, while bilateral touch, lateral line, auditory and bilateral vision in a second system. For zebrafish one study describes multiple paths as a “high road” through Hb (habenula, apical) and a “low road” through OT (optic tectum, bilateral) [do Camo Silva et al 2018].
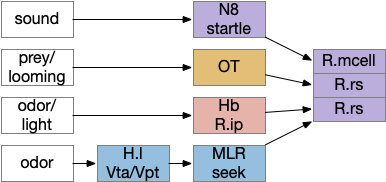
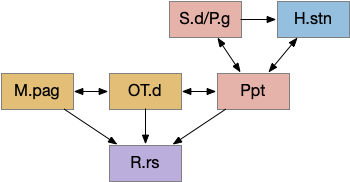
Some labeled paths for locomotion in vertebrates. H.l (lateral hypothalamus), Hb (habenula), MLR (midbrain locomotive region), N8 (acoustic-vestibular cranial nerve 8), OT (optic tectum), R.ip (interpeduncular nucleus), R.mcell (Mauthner-cell), R.rs (reticulospinal motor command)
The above diagram shows some vertebrate labeled paths, which is clearer in simpler vertebrates like the lamprey and zebrafish. In the zebrafish startle reflex, a sudden noise triggers a fast C-bend turn followed by rapid swimming. The trigger can be a noise, vestibular, or lateral line motion [Berg et al 2018]. The startle circuit is only three synapses from the original sensor to the muscle, from the N8 auditory/vestibular nerve to the giant M-cell (Mauthner cell in r4) to the motor neuron that drives locomotion. In young zebrafish larva, head touch neurons (N5 trigeminal) connect to M-cells and are later replaced by N8 [Kohashi et al 2012]. M-cells fire only once per escape to drive the initial turn. Interestingly the escape turn choice uses an axo-axonic repeater and amplifier [Guan et al 2021].
In a different path looming and dimming visual signals that represent predators or obstacles drive OT (optic tectum), which can drive escape that either uses or bypasses the M-cell depending on the threat level [Bhattacharya et al 2017]. OT also pre-programs the M-cell circuit by suppressing the left or right to avoid an obstacle [Zwaka et al 2022].
Phototaxis (seeking or avoiding light) uses a temporal gradient system composed of left Hb.m (medial habenula) and R.ip.d (dorsal interpeduncular nucleus), which projects to the R.rs (reticulospinal motor command) neurons via relays in V.mr (median raphe) and P.ldt (laterodorsal nuclei) [Chen and Engert 2014]. Food odor seeking uses the right Hb.m and R.ip.v (ventral interpeduncular nucleus) [Chen et al 2019].
In lamprey a distinct food-seeking path through V.pt (posterior tuberculum – possibly homologous to vertebrate Vta/Snc) to MLR (midbrain locomotor region) and finally to R.rs [Derjean et al 2010]. Zebrafish has a similar dual path through Hb.l (lateral habenula) through a midbrain TSN circuit [Koide et al 2018].
Slower escape uses a distinct prepontine (rhombomere r0-r1) circuit, which is suppressed by the M-cell escape circuits [Marquart et al 2019].
Some of these paths have shared elements, particularly at the motor control like MLR, but the general pattern is multiple labeled paths for each behavior. The paths already mentioned don’t include more complex food-seeking paths through the basal ganglia and hypothalamus.
Multiple labeled paths immediately raises the difficulty of coordination. How does the system juggle priorities? Even the simple startle reflex needs to be modulated because the animal shouldn’t startle if the loud sound is expected, such as near a waterfall. In contrast in a dangerous area with possible predators the animal should increase the reflex to a hair-trigger. Similarly if the threat is weak and the animal is hunting or eating and hungry, it might ignore the threat to continue eating. A second architecture, distinct from the label path, emphasizes the coordination of multiple paths, possibly using a consensus system to decide on an appropriate action.
Consensus loops
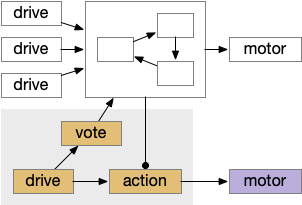
While the labeled paths have strong evidence, the consensus loop is only a thought experiment to tie the paths together. Multiple paths for food seeking and for avoiding is a distributed system, and distributed systems makes decision circuitry more complicated because they’re not central decision node. Every node needs to agree with the decision. Whether to avoid or approach needs to be agreed on by all the systems. It wouldn’t make sense for one system to believe the animal is approaching an object but another system believes the action is avoiding. Voting distributes the consensus.
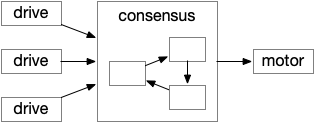
Illustration of a consensus loop. Multiple drives or labeled paths vote for approval to drive motor output.
The above diagram shows the model. The different labeled paths of seeking or avoiding join a voting consensus system in a motivational look, which allows one path to drive motor output.
Consensus system showing one path. The driving sense or motivation tries to disinhibit itself by voting in the consensus loop.
A single labeled path has a sense or motivation drive that tries to act on motor output, but is inhibited by the consensus system. For example, if a predator odor arrives, the odor avoidance path votes to enable its own locomotion. If the consensus system agrees, it will disinhibit the odor avoidance path, letting the animal escape. Note that a high priority threat could bypass the consensus system.
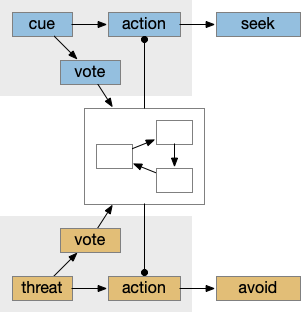
Seek and avoid consensus system
This system can manage conflicts between seek and avoidance, such as animals continuing to eat if a predator threat exists but is low. Consider a simplified consensus system with only one seek node and one avoid node, using the consensus to select one when there’s a conflict.
Managing conflicts between seeking and avoiding.
If there’s a food cue and no conflicting threats, the food vote passes easily and the animal seeks the food. Similarly a predator odor with no conflict will enable avoidance. If there’s a conflict, the system can weigh the costs and benefits of the threat and the food, possibly depending on hunger state or a more sophisticated threat assessment.
Keeping these general ideas of the labeled path and consensus systems in mind, let’s start working through several specific paths. The end goal is to organize the main brainstem locomotive areas into a simplified, unified model. The two major paths will be apical paths through Hb (habenula) using temporal gradients (klinotaxis) [Chen and Engert 2014] and bilateral paths through OT (optic tectum) using spatial gradients (tropotaxis).
Apical and bilateral avoidance
Because there are many labeled paths, dividing them up might help organize the model. An early division between labeled paths goes back to the bilaterian (worm-like, slug-like) ancestors, which added bilateral, dual-sensory navigation (tropotaxis, spatial gradient) to an older single-sensor navigation that used the animal’s movement to choose a direction (klinotaxis, temporal gradient), such as the simple tumble-and-run that even bacteria and simple radial zooplankton use for seeking odors (chemotaxis) and seeking or avoiding light (phototaxis). This chimera hypothesis [Tosches and Arendt 2013] considers bilateral animals as a fusion between the locomotive systems. The apical zooplankton larvae of bilaterian worms may have been a secondary development to escape predation [Mallatt 2021]. In vertebrates, apical klinotaxis is implemented by Hb (habenula) temporal gradient seeking and H.l (lateral hypothalamus) motivation. Bilateral tropotaxis navigation is implemented by several labeled path systems, typified by OT (optic tectum) and the M-cell start reflex.
A primitive apical example is the helical phototaxis of many annelid (marine worm) zooplankton larvae, and a primitive bilateral example is the mollusk sea slug navigation.
Zooplankton apical phototaxis
One type of zooplankton is essentially a globe with a fringe of cilia and an apical tuft for chemical processing, such as the Platynereis larva.
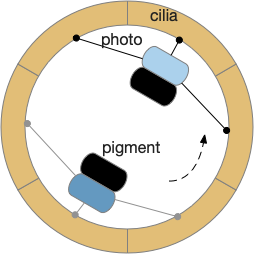
Phototaxis for this larva depends on its helical movement (helical klinotaxis). As it moves forward, the larva also rotates and wobbles, which means that parts of the equatorial band are nearer the light or further from the light depending on the rotation. If the upper cilia halt, the larva will steer toward the light. If the lower cilia halt, the larva will steer away from the light [Randel and Jékely 2016].
Phototaxis for an zooplankton larva.
The system depends on a directional eye, which uses a photoreceptor and a pigment cell that imposes directionality by shading the photoreceptor, because other cells of the larva are transparent. The photoreceptor compares the current brightness to its average brightness as the larva rotates. If it’s brighter than average, then it must be facing the light, and will signal the cilia to briefly halt, using ACh (acetylcholine) as a neurotransmitter. This trivial one-neuron circuit is sufficient for simple phototaxis [Randel and Jékely 2016].
Although this example larva uses two photoreceptors, it’s not truly bilateral and the two photoreceptors don’t communicate. Ablating one photoreceptor doesn’t abolish phototaxis, although it does reduce efficiency. Using three or four photoreceptor/pigment pairs would work, as well as removing all but one. This system is apical klinotaxis, not bilateral tropotaxis, which makes sense because the above zooplankton is not bilateral. While this zooplankton uses helical klinotaxis, another common form of klinotaxis is a side to side “casting” motion used by other simple animals like c. Elegans [Izquierdo and Lockery 2010].
If zooplankton phototaxis is an example of apical navigation, then the mollusk sea slug is an example of bilateral navigation.
Mollusk sea slug seek and avoid
The mollusk sea slug circuit is a pure bilateral circuit, almost directly a Braitenberg circuit [Braitenberg 1984], discussed in essay 14. The following shows a rough schematic of the sea slug seek and avoid. This circuit is interesting because with only a few neurons, the slug can switch from turning toward a food odor when hungry to turning away from the odor when not hungry [Gillette and Brown 2015].
Odor seek and avoid circuit for a sea slug. Hunger switches a food odor from seek to avoid.
In the diagram above, the central grey area is a switchboard circuit. Hunger reconfigures the switches connecting the odor to the turn motor neurons. When the slug is hungry, the right odor sensor connects with the left turn muscle, seeking the odor. But when the slug is sated, the right odor sensor connects with the right turn muscle, avoiding the odor. When the slug is hungry, it approaches food but when it’s not hungry, it avoids food odor cues.
A similar animal with a different circuit configuration uses serotonin to switch from avoidance to approach [Hirayama et al 2014].
For the goal of this essay, avoiding a failed food cue, this circuit is perfect because when the animal finds a false cue, it reversed movement from seek to avoid, which exactly fits the essay needs. Unfortunately, the vertebrate circuits aren’t nearly as straightforward. As a start for the vertebrate navigation paths, the startle reflex managed in vertebrates by the giant Mauthner cells is a simple starting point.
Amphioxus fast twitch reflex
The fast twitch startle reflex is a clear example of a bilateral labeled path avoidance circuit. A noxious sense on one side causes a fast turn away from the sense. The sense can be a touch on the head, such as running into an object, or a loud sound or a vestibular imbalance signal. This circuit predates vertebrates and a similar circuit exists in amphioxus, a filter-feeding chordate that looks something like a fish without a distinct head and without eyes, but with several photoreceptors including a frontal “eye.”
In amphioxus the startle reflex drives fast twitch muscle fibers, where normal swimming uses slow twitch fibers [Lacalli and Candiani 2017]. This circuit path is entirely distinct even to using the different muscles. The following diagram shows part of the amphioxus motor control circuit. (Because the neuron names are specific to amphioxus, they’re not hugely important for this essay.)
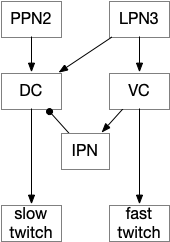
Amphioxus fast twitch escape uses LPN3, glutamate large paired neuron.
The diagram shows the LPN3 (large paired neuron) fast twitch escape path, and the PPN2 normal swimming match, including intermediary motor control neurons [Lacalli and Candiani 2017]. This amphioxus escape circuit resembles the zebrafish Mauthner cell escape.
Zebrafish Mauthner cell escape
Zebrafish have a pair of large M-cell (Mauthner cell) neurons that are specialized for auditory and vestibular startle escape. These are very fast reflexes on the order of 10ms, which can be modulated by higher context [Zwaka et al 2014] including OT. Although the M-cells perform a similar role to the amphioxus LPN3, it’s not clear that they’re homologous, which requires common descent, because the large escape neuron is a common pattern in non-chordate systems.
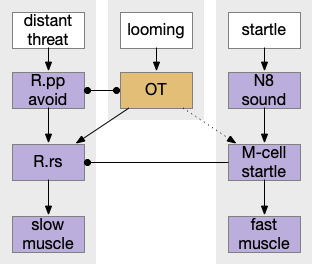
Zebrafish startle response at right in context with other labeled paths. M-cell (Mauthner R.rs cells in r4), N8 (acoustic/vestibular cranial nerve 8), OT (optic tectum), R.pp (prepontine avoidance in r0-r1), R.rs (reticulospinal motor control)
The primary input to M-cell escape is an auditory and vestibular signal from N8 (8th cranial nerve is auditory and vestibular). In water, sound and primitive vestibular sense have some similarities, because water motion produces not just sound but animal motion, depending on the frequency. The M-cell directly connects to motor neurons to muscles. The startle escape is only a three neurons and a clear, distinct labeled path.
A second zebrafish threat avoidance path uses neurons in R.pp (pre-pontine r0-r1) [Marquart et al 2019] for more distance threats. Unlike the M-cell circuit, this R.pp path is more than a reflex, but it’s still a hardwired path. A third threat circuit uses OT (optic tectum), for example the looming response. Most vertebrates will flee or freeze from a rapid and overhead expanding dark object, representing a potential predator or an obstacle. The mammalian startle circuit shares similarity with an acoustic projection to R.pn.c (caudal pontine reticular) neurons, in an analogous area to the M-cell [Kim et al 2017].
Some of these circuits do share sub circuits. For example, hindbrain locomotion and turning are distinct circuits that are used by both bilateral and apical avoidance circuits.
Hindbrain locomotion and turning
Senses are not the only source of distinct paths because actions can be split into parts like a car’s divided steering and acceleration. In vertebrates, accelerating and turning use distinct hindbrain circuits. Although both MLR (midbrain locomotive region) and OT.d (deep layer of the optic tectum) encode seeking and avoiding, they don’t encode left or right turns. Activating the left or the right MLR produces straight movement [Brocard et al 2010]. Turning is managed from OT.i (intermediate layer of the optic tectum) to distinct R.rs motor command neurons, marked by the chx10 transcription factory [Cregg et al 2020].
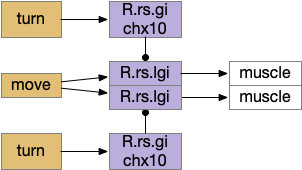
Hindbrain acceleration and turning circuits. R.rs (reticulospinal motor control)
The above diagram shows the basic idea. The upstream MLR can command forward movement without specifying details, because swimming is an oscillatory process with CPG (central pattern generators) in the spinal cord and the hindbrain. To turn, the chx10 neurons inhibit the swimming stroke in one direction [Cregg et al 2020], similar functionally to the apical zooplankton inhibition of cilia for phototaxis.
Splitting out turning can simplify the system by dividing labor, where OT.i is always responsible for obstacle avoidance, but a diverse set of labeled paths decode whether to seek or to avoid.
Optic tectum and dimming
The OT is named after its retinotopic visual map that is used for avoiding looming/dimming obstacles and predators, and also for seeking prey [Basso et al 2021]. For most vertebrates, OT is the primary visual area, and the visual cortex only provides abstract context, and amphibians and fish lack a proper visual cortex [Heap et al 2018]. For this essay, OT is less important for its sophisticated visual organization, but more because it also contains motor maps for seeking or prey and avoidance of looming objects, and dimming fields. Its motor map also contains drinking and licking [Liu et al 2022].
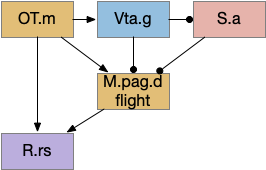
Looming/dimming path through optic tectum. OT.m (medial, deep optic tectum), R.rs (reticulospinal motor command)
OT processes looming and dimming objects and avoids them. Since the essay’s model lacks proper vision, the dimming is currently most important. Because OT also has obstacle avoidance, it’s a more sophisticated system than simply reflex. It’s likely that other avoidance systems will use OT for obstacle handling. Even in the case of the M-cell reflex, the OT.i pre-programs the M-cell, to avoid obstacles in case of a future startle [Zwaka et al 2014].
Optic tectum and turning
This division between turning and acceleration applies to OT itself. OT is a layered structure where the top layer is a visual map, the intermediate layer integrates other senses and produces turns, and the deepest layer includes actions such as avoiding and seeking [Liu et al 2022]. OT.d (deep OT) is a motor area for seek and avoid, connected with MLR and M.pag (periaqueductal grey) motor output, and integrates general dimming from the retina with distinct expansion calculation in OT itself [Heap et al 2018], to avoid looming objects. OT.i (intermediate OT) includes multi sensory integration and turning motor area, connected with LL (lateral line) electro sensation and water motion, somatosensory (whiskers in mice), auditory input from M.ic (inferior colliculus) and optic input from OT.s (superficial OT).
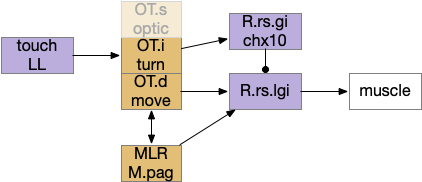
Optic tectum layered structure, emphasizing turning and motion. LL (lateral line water motion), MLR (midbrain locomotor region), OT.s (superficial optic tectum), OT.i (intermediate OT), OT.d (deep OT), R.rs (reticulospinal motor command)
Because only the top layer is specifically optic, some neuroscientists use “tectum” (roof in latin) instead of OT to emphasize its multi sensory and motor function, not just the optic features. On argument suggests that the optic layer OT.s is a secondary layer, added to a more primitive OT.i and OT.d that are more connected with reticular areas like MLR and M.pag [Edwards 1980], [Basso et al 2021]. With that argument, OT is primarily a moving and turning structure, receiving turning and moving input from touch, lateral line, primitive dimming, and other directional senses and combining with seek and avoid decisions. When the visual system developed enough detail to support crude images like looming disks or moving prey-like dots, the OT integrated vision into its top layer.
On the other hand, since OT.d receives dimming information from H.lg (central lateral geniculate nucleus) for looming escape [Heap et al 2018], it’s also conceivable that the base OT function is visual escape from dimming, where the later expanding, looming visual processing is an optimization.
Optic tectum obstacle avoidance combined with MLR seek or avoid movement. MLR (midbrain locomotor region), OT.i (intermediate optic tectum), R.rs (reticulospinal motor neurons).
This separation of obstacle avoidance turning from seeking and avoiding greatly simplifies some other circuitry that doesn’t need to duplicate the obstacle avoidance. Since other circuitry from the apical path, like the Hb-R.ip (habenula – interpeduncular nucleus) has its own turning system, OT.i doesn’t have a monopoly on turning. But even in that case, OT.i obstacle avoidance can inform apical navigation.
Some of these avoidance circuits are from the bilateral part of the chimera, such as the M-cell and the looming OT circuits, and others are from the apical part, such as Hb.m phototaxis, chemotaxis, and thermotaxis. So, let’s now more from the bilateral avoidance circuits, explore the vertebrate apical navigation.
Tunicate helical swimming and phototaxis
Tunicates (including sea squirts) are the closest chordates to the vertebrates, but because they have evolved at a greater rate and in specialized directions, comparison with vertebrates is difficult [Stolfi and Brown 2016]. Ascidian tunicates (sea squirts) have a mobile tadpole stage that plants itself in under 24 hours and transforms into a sessile filter feeder, reforming the entire brain. In general, neuroscientists believe amphioxus more resembles the ancestral vertebrate, and that ascidians have lost too many ancestral structures for a reasonable comparison [Holland 2016]. But for the sake of exploration let’s run through a thought experiment as if the ascidian larva is a compressed and simplified version of the vertebrate ancestor, although possibly only the vertebrate larva.
Specifically, consider phototaxis in the apical helical klinotaxis mode that follows a temporal gradient, since both amphioxus and ascidian larva swim in a helical pattern. Even bacteria can follow odor gradients [Hengenius et al 2012] and as discussed above zooplankton phototaxis can move toward light with only a single photosensor [Randel and Jekely 2016]. Both amphioxus and ascidian larva have single unpaired eyes, amphioxus as a single frontal eye [Lacalli 2022] and ascidians with an asymmetrical eye paired with a second pigment cell used for geotaxis as a primitive vestibular sense [Hoyer et al 2024]. In both cases, the “eye” is directional with a pigment cell, but a non-image-forming collection of photoreceptors. The ascidian asymmetrical eye works because the ascidian tadpole swims in a helical pattern so the timing of the light on the eye matters more than its position [Ryan et al 2016].
Ascidian larvae swim in a helical pattern comprised of unilateral tail flicks and symmetrical swimming [Ryan et al 2017] and use the asymmetry of the photoreceptor and photopigment to swim toward light [Mast 1921], [Zega et al 2006]. Since helical swimming doesn’t need stabilizing fins or vestibular systems to manage roll, yaw, and pitch with 3d swimming, it’s available to evolutionarily simpler systems. Another advantage of helical phototaxis is that the photoreceptors are auto-calibrating by simply averaging the light in a rotation and requires less circuitry than a bilateral comparison of light [Randel and Jekely 2016].
However, unlike the trivial zooplankton circuit that directly connected the photoreceptor to arrest the cilia, ascidian larvae need to modulate the bilateral swimming in the primitive hindbrain, timing the muscle inhibition to achieve the same effect.
The ascidian ocellus (“eye”) has two types of photoreceptors with distinct responses. Type 1 has a pigment and lens and is directional (37 cells), while type 2 is non-directional (no pigment partner) [Salas et al 2018]. If the pigment is genetically deleted, the animal can’t use phototaxis but does respond to dimming with an escape response. In other words, the dimming response and phototaxis use distinct labeled paths with distinct input neurons [Kourakis et al 2019]. The following shows the circuit for the type 1 photoreceptors for phototaxis, where the boxes represent single neurons or small collections (5-8) of neurons, not large functions (from [Ryan et al 2016]).
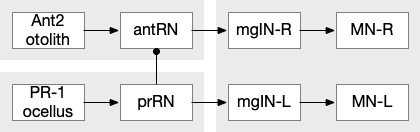
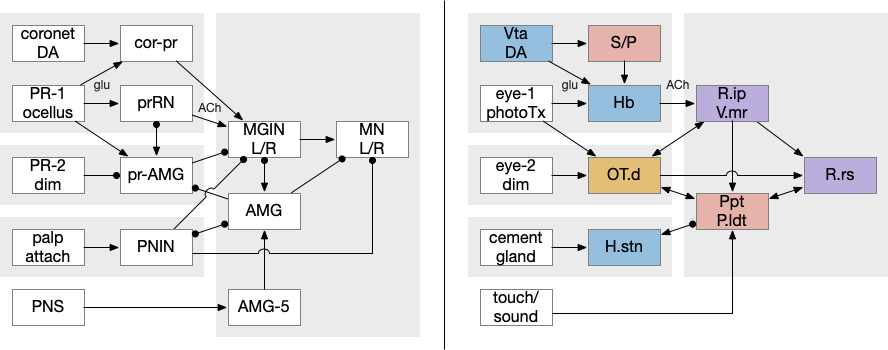
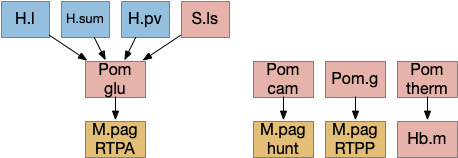
Ascidian larva phototaxis (ocellus) and geotaxis (otolith) circuit. Ant2 (antenna geotaxis sensors), antRN (antenna relay neuron), mgIN (motor ganglion interneurons, left and right), MN (motor neurons, left and right), PR-1 (type-1 phototaxis photoreceptors), prRN (photoreceptor relay neuron)
The above diagram shows both geotaxis and phototaxis circuits, which are specific to right or left motor neurons respectively, because the ascidian larva neuron circuits are highly asymmetrical. Ascidian larva geotaxis swims upward and phototaxis swims away from light, generally downward. The combination encourages swimming to the underside of ledges, such as the underside of boats and harbor piers [Ryan et al 2016]. Because of the helical swimming, the left and right motor neurons aren’t left or right turns, but turns toward or away from the target. Although this circuit is more complicated than the purely apical zooplankton because of the interface to bilateral swimming, the helical swimming keeps the circuit relatively simple.
The above partial circuits are complicated by the coronet cells, another sensory cell that are paired with the photoreceptors, but with unknown function. The circuit connectivity is interesting, because coronet cells modulate both the phototaxis and geotaxis paths, but aren’t a path of their own. The phototaxis and geotaxis relay neurons above are partially bilateral. Only 70% of their connectivity is to the main side, but 30% of the connectivity is to the opposite side. In contract, the coronet-enabled neurons are 100% to the main connection [Ryan et al 2016].
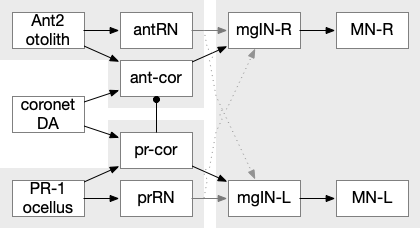
Coronet cell modulation of phototaxis and geotaxis in the ascidian larva. ant2 (antenna geotaxis cell), ant-core (antenna-coronet relay neuron), antRN (antenna relay neuron), DA (dopamine), mgIN (motor ganglia interneuron, left and right), MN (motor neuron, left and right), PR-1 (photoreceptors type-1), pr-cor (photoreceptor-coronet relay neuron), prRN (photoreceptor relay neuron)
As a thought experiment (unsupported by scientific evidence) the main phototaxis path might be uncertain and stochastic, while the coronet-enabled path would be a certain, deterministic connection. If the coronet cells measured the certainty of the animal’s current direction, it could encourage sticking to the current path. For example, if the coronet cells were food-odor gradient sensors, they could fire when the animal was heading toward food, enabling a chemotaxis based on modulation of geotaxis and phototaxis.
Tunicate dimming response
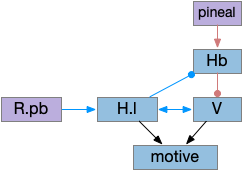
The ascidian dimming response triggers locomotion with a strong turn as an escape response to predators [Kourakis et al 2019]. Unlike the phototaxis photoreceptor, the dimming photoreceptors are non-directional because’er not shaded by the pigment cell. There are 23 type-1 directional photoreceptors and 7 type-2 non-directional photoreceptors for dimming.
The above diagram shows the dimming path in context with the previous phototaxis path. Like the phototaxis path, the dimming path starts from the type-2 photoreceptors to a relay neuron and to the control neurons in the motor ganglion. Unlike the phototaxis path, the dimming path is modulated by ascending motor signals from AMG (ascending motor ganglion) and from the phototaxis path [Ryan et al 2016], presumably so the normal helical phototaxis doesn’t trigger a dimming response.
Cement gland and attachment
The ascidian larva hatches before dawn, swims upward for a few hours because geotaxis is enabled before phototaxis neurons attach, and then swims away from the light, settling on a lively rock, preferring a ledge to settle under if possible. Larva do not feed [Ryan et al 2016]. The larva will attach with a cement gland on the front of its head, a trio of palms, and then transforms into the adult sessile filter feeder. The palp sensors trigger the attachment circuit, which stops all swimming and begins the metamorphosis [Anselmi et al 2024].
Although the full details of the above circuit [Ryan et al 2016] aren’t critical, the PN (palp neuron) senses the animal bumping into a rock modulated by chemical senses that avoid toxic area, and triggers a swimming shutdown by inhibiting the motor neurons and interneurons [Hoyer et al 2024]. Like the previous diagrams, the boxes represent individual neurons or small group, not large functional regions.
While the ascidian cement gland is permanent, several fish [Pottin et al 2010] and amphibians [Rétaux and Pottin 2011] have a homologous cement gland used for larvae, not adults. For example, frog tadpoles can attach to the bottom of leaves or to the water surface to avoid predators until they are large enough to hunt [Jamieson et al 2000], [Yoshizawa et al 2008]. Because of the widespread cement gland among many fish species and amphibians as well as the tunicates, it’s likely the original vertebrates had a similar cement gland [Rétaux and Pottin 2011]. Whether the gland was larva-only like in vertebrates or also used for adults as in tunicates is unknown. In either case, the cement gland circuit that inhibits locomotion must have been part of the original vertebrate.
Vertebrate analogies to the ascidian circuits
Because the ascidians are so specialized and reduced from the common ancestor with vertebrates, including major losses in genes, cells and structures, comparing the two is essentially impossible to be homologous (shared descent) [Holland 2016]. However, for the sake of exploration, I’m ignoring that advice, and looking for analogous vertebrate circuits to the ascidian larva.
The ascidian behavior each have distinct circuit paths that mostly only come together at the motor control neurons. The exception is the feedback from the AMG (ascending motor ganglion) neurons, which do feedback to the midbrain neurons, but the main paths are separate forward paths. Each of the geotaxis, phototaxis, dimming, cement gland attachment, and bilateral escape are circuit paths that are distinct until the motor command neurons.
A vertebrate analogy to the ascidian phototaxis gradient path might be the path from the retina to Hb.m (medial habenula) to R.ip (interpeduncular nucleus) and V.mr (median raphe), which then project to R.rs (reticulospinal motor command). Like the ascidian path, the Hb-R.ip phototaxis path is relatively isolated from the other paths, although Hb.m does receive large modulation from the hypothalamus. Although R.ip is mostly descending, like ascidian mgIN, V.mr is both ascending and descending like mgIN and AMG.
The dimming path from the type-2 photoreceptors resembles the dimming input to the vertebrate OT (optic tectum). Although existing vertebrates have more sophisticated eyes that can distinguish expanding objects, the dimming input to OT is still important and used for escape directionality [Fotowat and Engert 2023], [Heap et al 2018]. Retina dimming cells reach AF6 and AF8 [Temizer et al 2015], which are thalamic arborization fields before reaching OT.d. Although more complicated expanding looming response in vertebrates is better studied, expansion detection requires an image-supporting eye, and OT.d receives the simpler dimming input. Like the ascidian dimming pr-AMG (photoreceptor – ascending motor ganglion) neuron, OT.d receives multiple ascending and descending inputs that modulate the dimming response. In particular Ppt (pedunculopontine nucleus) and P.ldt (laterodorsal nucleus) both receive OT.d output and forward to R.rs, functionally similar to mgIN (motor ganglion interneuron), and send ascending feedback from R.rs to OT.d, resembling the AMG (ascending motor ganglion) functionality.
Because the cement gland exists in vertebrates, the circuit should be available, and studies do show that N5 (head touch trigeminal nerve) innervates it automatically [Pottin et al 2010], but I haven’t read any study that says this this specific group of trigeminal neurons connects to. As a through experiment, consider H.stn (subthalamic nucleus) as a choice for the cement gland, because H.stn halts ongoing action, and because H.stn receives direct input from C.i (insular cortex) and C.ss (somatosensory cortex), which are more sophisticated versions of the chemo / mechanosensory palp neurons.
The coronet path enhances taxis confidence, reducing stochastic choice, and is a set of dopamine neurons. The striatum circuit and dopamine’s role has a similar function. Without dopamine, the basal ganglia suppress weak input, and allow stochastic action. With dopamine, the basal ganglia suppresses the randomness and keep action on track. This path resembles rheotaxis food seeking, where a fish approaches a food odor by swimming upstream [Coombs et al 2020]. The “what” signal (odor) differs from the “how” signal (water current cues). Like rheotaxis, the coronet cells enhance the existing phototaxis and geotaxis, reducing the default stochastic noise.
Hb.m Medial habenula
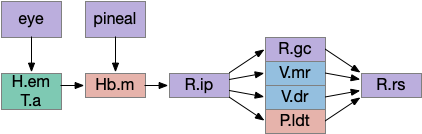
Of the ascidian labeled paths above, the Hb (habenula) phototaxis path will be a useful anchor for the upcoming consensus circuit. Like the ascidian asymmetrical phototaxis neurons, the vertebrates Hb.m (medial habenula) is also governed by Nodal asymmetry [Roussigne et al 2009], where Nodal is a developmental genetic transcription factor. In zebrafish the left Hb.m support phototaxis, and the right Hb.m supports chemotaxis [Chen et al 2019]. Hb.m phototaxis receives both “on” and “off” neurons from the retina with a relay either in H.em (pre thalamic eminence) [Zhang et al 2017] or T.a (an area in the anterior thalamus) [Cheng et al 2017], where the connections are debated. Although Hb.m does receive dimming input from the adjacent photoreceptive pineal gland, the retina photoreceptors are more important for phototaxis [Dreosti et al 2014].
As an anatomical note, the zebrafish Hb.m is actually dorsal and therefore named Hb.d. Similarly the zebrafish Hb.m is ventral and named Hb.v, but as a simplification I’ve used the mammalian name.
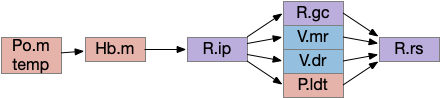
The output path from Hb.m is through R.ip (interpeduncular nucleus), which projects to several areas including R.gc (pontine central era), V.mr (median raphe – serotonin), V.dr (dorsal raphe – serotonin), and P.ldt (laterodorsal nucleus – ACh) [Quina et al 2017]. The V.mr glutamate and GABA neurons may be more important for this circuit than the serotonin neurons, which they outnumber. Also, note that V.mr is located in the same hindbrain rhombomeres (r2-r5) as some of R.rs, but are more ventral, and are reciprocally connected. In other words, V.mr is highly action and motor associated.
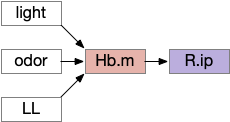
As described above, the Hb.m-R.ip path is a klinotaxis path for phototaxis [Chen and Engert 2014], chemotaxis and thermotaxis, where the klinotaxis is temporal from the animals movement, but not the helical movement of the ascidian larvae. The Hb.m-R.ip klinotaxis has multiple inputs for lamprey, including light, odor, and lateral line (water movement) [Stephenson-Jones et al 2011].
Habenula klinotaxis for lamprey for light, odor, and lateral line. Hb.m (medial habenula), LL (lateral line), R.ip (interpeduncular nucleus)
Although I’ve focused on Hb.m as an avoidance gradient circuit, it’s also a food odor seeking circuit [Chen et al 2019]. The Hb.m klinotaxis for light and odor also applies to temperature, using input from Po.m (medial preoptic nucleus) [Palieri et al 2024] and social seek and avoidance [Okamoto et al 2021], [Chou et al 2016].
Because Hb.m has several sub-nuclei and genetic clusters, it likely represents different labeled paths, supporting multiple distinct seek and avoidance paths. A binary seek vs avoid circuit is likely an oversimplification, because studies have found at least 5-6 olfactory Hb.m clusters in the larval zebrafish [Jetti et al 2014], [Beretta et al 2014]. Hb.m is asymmetrical, like the ascidian larva. Odors from either olfactory bulb activate the right Hb.m [Chen et al 2019]. Hb.m neurons have at least 262 neuropeptide receptors [Ables et al 2023] as well as morphine receptors [Gardon et al 2014], [Boulos et al 2020] including neuropeptides modulating hunger or social motivation from hypothalamic areas like H.l and H.pv.
R.ip interpeduncular nucleus
Since I’ve already covered some of the R.ip klinotaxis function in essay 24 and essay 25, I’m going to focus on the R.ip connectivity, particularly the ascending connectivity. R.ip descending efferents don’t target R.rs directly, but instead use intermediaries like R.gc (pontine central gray), V.mr (median raphe) and P.ldt (laterodorsal tegmental area) [Lima et al 2017], [Quina et al 2017].
The ascending afferents of R.ip also work through intermediaries, particularly P.ldt and V.mr [Quina et al 2017], although other connectivity studies report R.ip as directly producing ascending connectivity [Lima et al 2017]. Because V.mr is directly caudal to R.ip, the disagreement is essentially about the boundaries between R.ip and V.mr.
The ascending R.ip connectivity will become important in the next section on the consensus circuit because it completes the consensus loop, where other labeled path connectivity is descending. The ascending role is analogous to the ascidian AMG (ascending motor ganglion) neurons. For a consensus circuit to work, all nodes need to be informed of the consensus decision.
Consensus circuit narrative
Let’s now consider the consensus circuit and how it might develop from a strict labeled path system. This is just a thought experiment as a narrative explanation for the Hb.l (lateral habenula) system.
For simplicity, let’s restrict the narrative to apical systems only, ignoring bilateral systems like OT, and let’s start from a labeled path system. In the lamprey, odor information from Ob (olfactory bulb) splits into multiple paths. One path reaches Hb.m directly and another contacts V.pt (posterior tuberculum), considered a homologue of Vta / Snc (substantia nigra pars compacts), which then contacts MLR in lamprey [Derjean et al 2010] and zebrafish [Kermen et al 2013].
In this example, these two paths are distinct with threat odors going through the Hb.m – R.ip circuit and using P.ldt as an apical version of the MLR (which in lamprey may not be distinct from Ppt MLR, since the lamprey doesn’t have distinct Ppt, P.ldt and M.cnf (cuneiform nucleus)). The animal seeks food using the V.pt to MLR path.
These two system can come into conflict. For the above simple system, suppose conflicts are resolved in R.rs itself, as a hard-coded priority where threats always win. But now consider a system where the conflict is resolved earlier in the stream by adding Hb.l as a lateral inhibition relay.
As a first step consider lateral inhibition of threat odor suppressing food seeking. Here the lateral inhibition path uses a relay from Hb.m to Hb.l [Gouveia and Ibrahim 2022] in a primitive Hb.l that then suppresses the V.pt path. This lateral inhibition duplicates the earlier lateral inhibition in R.rs, but is more specific because it inhibits earlier in the two paths.
In the above diagram, the blue lines represent new connections. Notice the gating pattern for V.pt resembles the gating for the consensus circuit where the action nodes are V.pt and V.mr. Hb.l then becomes the vote accumulator for the consensus circuit. Also notice the similarity with the sleep model from essay 29, where Hb inhibits food seeking for sleep. An alternative narrative might repurpose the sleep inhibition into a path inhibition [Hikosaka 2010].
Both Hb.l and Hb.m are tonically acting, meaning that without any input their resting output is a middle value, not a binary output. This means Hb.l can gate V.pt seek and also gate its opposing avoidance circuit in V.mr and P.ldt.
For the next step, let’s add both a bidirectional selection and also add some internal state management, because the animal shouldn’t seek food if it’s sated.
H.l hunger modulation
H.l (lateral hypothalamus) has access to hunger and satiety information by sensing blood levels directly and from connections from R.pb (parabrachial), which has signals from the digestive system via N10 vagus nerve through R.nts (solitary tract nucleus). When Ob (olfactory bulb) senses a food odor , H.l can modulate it with the current hunger sense. This means H.l as gating input to Hb.l is more effective than the simple lateral inhibition from Hb.m If the animal is sufficiently hungry, it might ignore weak threats. Note the similarity to the mollusk sea hare circuit, where hunger changed food odor from seeking to avoidance depending on the internal state.
Adding H.l hunger modulation to the decision between the threat odor avoidance path and the food odor seek path. H.l (lateral hypothalamus), Hb.l (lateral habenula), Hb.m (medial habenula), MLR (midbrain locomotor region), P.ldt (laterodorsal tegmentum), R.ip (interpeduncular nucleus), R.rs (reticulospinal), V.mr (median raphe), V.pt (posterior tuberculum).
In addition to hunger, other internal states can modulate Hb such as hypothalamic threat signaling ([Wagle et al 2022]. This step also adds control of the threat path, taking advantage of the Hb.l tonic activity to either inhibit food seeking or threat avoidance.
Place avoidance without a threat
Suppose we take the above circuit, but ignore or disable the threat avoidance path via Hb.m. Even without the threat path, there is an avoidance path from Hb.l to V.mr and P.ldt, where Hb.l not only disinhibits threat avoidance, but can produce place avoidance without a threat.
The above diagram shows deletion of the threat path, while retaining the abstract place avoidance path. If place avoidance is triggered, the animal will avoid the current location without needing a specific threat to avoid. This means that H.l stimulation by itself can trigger real-time avoidance [Stamatakis et al 2016]. In mammals the H.l to Hb.l connection has at least 6 clusters [Calvigioni et al 2023], which suggests multiple paths even in the abstract place avoidance.
S.v ventral striatum digression
This model of the seek vs avoid circuit can be extended to S.v (ventral striatum aka nucleus accumbens) and P.v (ventral pallidum). Consider S.v / P.v as a generalization of H.l, providing more general context beyond hunger. This basal ganglia extension allows for a positive feedback loop. which enables multiple rounds of voting, integrating values, such as with drift diffusion.
In the above, I’ve split the V.pt of the lamper into an ascending Vta (ventral tegmental area) dopamine area from mammals, but left the V.pt to represent the descending glutamate / GABA portion of Vta, despite mammals lacking a distinct V.pt. If there’s a food cue when hungry, H.l to Vta stimulation will generate high DA in S.v, enabling it, which will disinhibit V.pt to enable food seeing. Here, S.v / P.v is acting as the consensus circuit and the V.pt path is the action for food seeking.
As with the smaller Hb.l circuit, S.v / P.v is also part of a sleep / wake circuit using dopamine as a wake signal, as used in essay 29. If the animal is currently seeking food, it shouldn’t fall asleep, and the high dopamine signals to stay away. Again, from a narrative sense, this circuit could have been repurposed from a wake circuit, as opposed to a path conflict system.
In zebrafish Hb.l only projects to V.mr and does not project to any DA [Amo et al 2014], while in the more primitive lamprey Hb.l projects to both V.mr and DA [Stephensen-Jones et al 2011], which suggests that the V.mr projection is more functionally critical to this circuit than the Vta projection, or that the Vta circuit is a later development. The zebrafish V.pt has descending dopamine but the existence of significant projections to the striatum is questioned [Yamamoto and Vernier 2011].
Note that H.l retains its central role, where the S.v circuit generalizes the base H.l function without replacing it. Stimulating H.l.g (H.l GABA neurons) can trigger seeking through its projection to Vta [Nieh et al 2016], and stimulating H.l.glu (glutamate H.l neurons) can trigger place avoidance through the H.l.glu projection to Hb.l [Stamatakis et al 2016].
Hippocampus digression
For place preference and place avoidance E.hc (hippocampus) plays a natural because E.hc represents context and place such as place cells, and H.hc projection strongly to both the hypothalamus and S.v. If we add the H.hc projections to H.l via S.ls, the seek / avoidance circuit looks something like the following.
Hippocampus modulation of H.l place seek and avoid. DA (dopamine), E.hc (hippocampus), H.l (lateral hypothalamus), Hb.l (lateral habenula), P.v (ventral pallidum), S.ls (lateral septum), S.v (ventral striatum), Vta (ventral tegmental area).
H.l has neurons that represent food zones and non-food zones [Jennings et al 2015], presumably using E.hc place information, although possibly using P.bst (bed nucleus of the stria terminals) as an intermediary.
H.sum completing consensus loop
The consensus circuits needs to return the final action and motor choice back into the early layers, otherwise the motivation circuit wouldn’t know if a lower-level startle or OT looming escape took priority of the seek path. With analogy to the ascidian larva, this role resembles the AMG (ascending motor ganglia) neurons, which I associated with P.ldt and V.mr. For this consensus narrative, I’m taking H.sum (supramammillary) as the primary feedback node with an assist from Poa (preoptic area) to complete the loop to Hb.m and M.pag (periaqueductal gray).
H.sum as completing the consensus loop, linking the habenula output back to habenula input. H.l (lateral habenula), H.sum (supramammillary nucleus), Hb.m (medial habenula), MLR (midbrain locomotor region), M.pag (periaqueductal gray), P.ldt (laterodorsal tegmentum), Poa (preoptic area), R.ip (interpeduncular nucleus), R.rs (reticulospinal), V.mr (median raphe), V.pt (posterior tuberculum).
H.sum has several sub circuits with different functions, which studies are only starting to untangle. H.sum tac1 (neurotransmitter aka substance P) is strongly associated with upcoming locomotion [Farrell et al 2021]. H.sum’s Poa projection is specifically associated with threat avoidant locomotion [Escobedo et al 2023].
V.mr and P.ldt are connected with R.rs and the bilateral OT circuit, and therefore have information about the selected action at the level of the hindbrain and motor afferent copies. Both are strongly connected to H.sum. H.sum also connects with M.pag (periaqueductal gray) and H.sum activates when M.pag.d is stimulated [Pan et al 2004]. H.sum also activates when the H.vm (ventromedial hypothalamus) threat nuclei are stimulated.
H.sum is immediately rostral to Vta and highly connected with it (not shown in the diagram.) H.sum contains some DA neurons itself, which are sometimes considered as an extension of A10, the Vta dopamine neuron area, although the neuron types differ [Yetnikoff et al 2014], [Menegas et al 2015].
H.sum is strongly connected with E.hc (hippocampus) and is one of the few external input to both E.dg (dentate gyrus) and E.ca2 (cornu ammonia), and is a major theta source to P.ms (medial septum), which drives E.hc theta. Its link to E.hc are important for both novel object exploration [Chen et al 2020], [Takahashi et al 2023] and social memory [Qin et al 2022]. Although I’m not yet adding E.hc to the essays, the novel object detection will be important soon to avoid repeated exploration of the same object.
Note that Poa has already participated in the Hb.m to R.ip circuit because Poa drives thermotaxis [Palieri et al 2024] as part of the original Hb aversive apical path.
M.pag tetrapod complications
In a sense, the vertebrate brain is designed around fish navigation, exemplified by the simple M-cell startle circuit that requires only three neurons between the acoustic sense and the swimming muscles. Although the direct Braitenberg-like connections to R.rs work for fish locomotion, tetrapod locomotion is more complex. M.pag (periaqueductal grey) is a central grey area surrounding the midbrain ventricle (“periaqueductal”), and it an inner ring to OT, which is immediately dorsal to it. Naming it “OT.dd” (deep, deep layer of OT) would not be unreasonable. Among other tasks like vocalization [Jürgens 1994] and hunting [Marín-Blasco et al 2020], M.pag provides a similar to R.rs but at a higher level, like syllables to phonemes. So in the following examples, M.pag can be viewed as similar functionality to R.rs.
Unlike R.rs, M.pag can access more sophisticated navigation. Where the M-cell can only turn left or right, M.pag can use OT for obstacle avoidance and even higher navigation of the hippocampus using H.pm.d (dorsal premammillary nucleus) [Wang et al 2021].
M.pag flight
M.pag implements innate behaviors, including flight, freezing, hunting, grooming, and vocalizations. The following diagram shows some of the looming flight circuitry [Zhou et al 2019]. As before, OT.m primarily processes the looming signal and OT.m sends input to M.pag.d as an integrated threat signal, where M.pag.d computes a threshold for responding to the threat [Evans et al 2018].
M.pag flight for the looming circuit. M.pag.d (dorsal periaqueductal gray), OT.m (medial, deep optic tectum), R.rs (reticulospinal), S.a (central amygdala), Vta.g (gaba neurons of the ventral tegmental area).
In the diagram, the second interesting path is through Vta.g (Vta GABA neurons) and S.a (central amygdala). Because OT.m and M.pag.d directly output to R.rs neurons, the projects to Vta.g and S.a aren’t required for motor control, but because of the distributed consensus system, other systems need to be informed of the looming response. S.a modulates defense, hunting, and eating systems, and Vta.g also inhibits the current action by suppressing dopamine, back to the consensus loop, suppressing any current seek action.
M.pag.vl avoidance
While M.pag.d is strongly associated with fast escape, M.pag.vl is more complicated with diverse functions including hunting [Franklin 2019], [Marín-Blasco et al 2020], vocalization [González-García et al 2024], and laughter [Klingbeil et al 2021]. Since this essay focuses on avoidance, where avoidance here isn’t the high speed predator escape of M.pag.d.
As discussed above, H.l is a central motivational node, filling a similar role to the central hunger node in the mollusk sea hare navigation. However, H.l is much more complicated than a simple hunger node. One developmental paper divided H.l into nine distinct regions [Diaz et al 2013], but that anatomical division understates the complexity. A genetic transcription analysis finds 15 glutamate and 15 GABA clusters [Mickelson et al 2019]. Interestingly, the Diaz study identifies their H.l.1 area with H.sum.l, treating H.sum.l as part of H.l.
In general, H.l.glu produces place avoidance and H.l.g enables seeking, but as mentioned above with at least 15 genetic types and 9 regions, this division is almost certainly an oversimplification.
H.l seek and avoid efferents. E.ca1.v (ventral hippocampus), H.l (lateral hypothalamus glutamate and GABA), Hb.l (lateral habenula), M.pag (periaqueductal gray), S.ls (lateral septum), Vta.g (ventral tegmental area GABA).
The H.l.glu to M.pag connection is certainly capable of driving motor avoidance. Interestingly, a different H.l population is part of the M.pag hunting circuit. Both Vta.g and Hb.l enter the motivation loop. I’ve added the E.ca1.v (ventral hippocampus CA1) input to H.l because E.hc.v (ventral hippocampus) is strongly associated with place, and E.hc.v specifically with aversive context.
R.pb peribrachial nucleus
R.pb (peribrachial nucleus) is a pain, alarm, feeding, and respiration hub in the prepontine isthmus area (r0-r1). As an alarm center [Campos et al 2018], R.pb is connected with escaping and avoiding circuits. As covered in essay 29 speed, it includes a high Co2 trigger that drives place avoidance. It also includes pain triggers for escape. R.pb has multiple functions defined more by chemical markers than topology. One study explored R.pb’s role in escape and avoidance behavior [Chiang et al 2020].
R.pb avoidance circuits. dyn (dynorphin neurotransmitter), H.vm (ventromedial hypothalamus), M.pag.l (periaqueductal gray), P.bst (bed nucleus of the stria terminalis), R.pb (peribrachial nucleus), RTPA (real-time place avoidance), S.a (central amygdala), tac1 (tachykinin 1 / substance P neurotransmitter)
R.pb.dl (dorsolateral R.pb) and R.pb.el are adjacent R.pb areas that are associated with alarm and pain responses. R.pb.dl receives direct N5 (trigeminal – head, jaw) and N.sp (spinal) pain input, including pain input marked by tac1 (tachykinin 1 peptide aka substance P). Relevant to this essay, the outputs divide between direct escape behavior with not learning and indirect avoidance behavior with learning. The M.pag.l projection produces flight and jumping. The S.a (central amygdala) and P.bst (bed nucleus of the stria terminalis – extended amygdala) projections produce real-time place avoidance and are capable of CPA (conditioned place avoidance) [Chiang et al 2020]. The R.pb example is useful because it combines a direct locomotion to M.pag with output to the slower consensus circuit.
Preoptic area
Poa (preoptic area) is a multifunctional area directly anterior to the hypothalamus and often considered part of the hypothalamus, although genetic markers suggest it’s more closely related to the forebrain. Like other brainstem areas, its functionality is more organized by genetic markers than topology.
The above diagram shows some of the Pom (medial preoptic area)functions. Temperature management has been discussed with a connection through Hb.m gradient following. Threat avoidance from signals from H.sum, H.pv (periventricular hypothalamus), or S.ls (lateral septum) can lead to RTPA through a M.pag projection [Escobedo et al 2023]. Local exploration, a RTPP function, also uses a M.pag projection [Shin et al 2023], and Pom can also enable hunting [Park et al 2018], although through a M.pag projection. The recent genetic research tools will likely unravel more of its functionality.
Poa has a strong projection to both Hb.m and Hb.l, suggesting that it’s an important node in the locomotion consensus circuit. In the thought experiment I’ve outlined above, Poa is part of the feedback system through H.sum, but Poa also receives E.hc.v (ventral hippocampus) input through S.ls (lateral septum), so it may be an important node in its own right.
Ppt / P.ldt
The ACh (acetylcholine) neurons near the midbrain-hindbrain boundary Ppt (pedunculopontine tegmentum) and P.ldt (laterodorsal tegmentum) are the core of the MLR. In simpler vertebrates like the lamprey, the MLR is only a single area, generally named P.ldt. In mammals, not only are P.ldt and Ppt split, but a chunk of locomotive action is in a different nucleus M.cnf (cuneiform). Although M.cnf is more of a direct locomotive area, the locomotive neurons don’t respect the anatomical boundary, but are a group of glutamate neurons spanning from Ppt to M.ncf, where Ppt and M.cnf are neighbors [Caggiano et al 2018]. Tetrapod locomotion is more complex than fish swimming, which may be a partial reason for the expansion and division.
Ppt is strongly reciprocally connected with the deeper layers of OT: OT.i for turning and sensory integration, and OT.d for seek and avoid. Its connections resemble the R.pgb (parabigeminal aka nucleus isthmi) which sustains attention for the OT.s (superficial OT) [Knudsen 2011] and covered in essay 19. R.pgb, Ppt, and P.ldt are sibling nuclei that develop from the same area in r1 that also produces R.pb and cerebellum granule cells [Pose-Méndez et al 2023].
Some P.ldt connections, emphasizing that Vta connections are collaterals of R.rs. H.sum (supramammillary), P.ldt (laterodorsal tegmentum), R.rs (reticulospinal), Vta (ventral tegmental area)
P.ldt is complicated by the relative lack of recent studies of its descending projections since [Cornwall 1990] and an over-focus on its Vta connection. Because neuron tracing in [Zhao et al 2023] suggests that P.ldt has more descending connections to R.rs than Ppt and that all Vta connections are collaterals of R.rs connections, studies like [Coimbra et al 2021] and [Liu et al 2022] that find locomotion through Vta projections could be produced by its R.rs projection. P.ldt has reciprocal connections with H.sum.
Vta
Although Vta (ventral tegmental area) is most studied for its ascending dopamine projections to S.v (ventral stratum) and F.pfc (prefrontal cortex), it also contains glutamate and GABA projections, including descending connections. Non-tetrapods like fish and lamprey have a homologous V.pt (posterior tuberculum) with prominent descending locomotor connections to MLR [Ryczko et al 2017], [Derjean et al 2010]. The earlier thought experiment for the development of a locomotor consensus split out an ancient V.pt from the mammalian Vta as a way of describing the old descending functionality.
Vta glutamate and GABA connections. H.l.glu (lateral hypothalamus glutamate), Hb.l (lateral habenula), M.pag (periaqueductal gray), OT (optic tectum), P.bst (bed nucleus of the stria terminalis), S.a (central amygdala), S.am (medial central amygdala), S.msh.pv (medial shell of the ventral striatum, parvalbumin neurons), Vta.da (ventral tegmental area, dopamine), Vta.g (Vta GABA), Vta.glu (Vta glutamate)
The above diagram shows some of the connections of the glutamate and GABA Vta [Taylor et al 2014], including projections to M.pag and to Hb.l that are direct locomotor for seek and avoid. The Vta is a main dopamine source for S.v and F.pfc with multiple distinct areas. Vta.m, which projects to S.msh (medial shell of S.v) is aversive, while Vta.l, which projects to S.lsh (lateral shell of S.v) and S.core (core of S.v) promotes seek [Szőnyi et al 2019]. Vta.m is non-reinforcing, as opposed to Vta.l, which is well-studied for reinforcement.
P.v ventral pallidum
P.v is a main output of S.v and the only output of S.ot (olfactory tubercle). As essay 29 covered, it’s an important sleep/wake node. For this essay, the important bit is a split between RTPP and RTPA depending on its output.
Calvigioni D, Fuzik J, Le Merre P, Slashcheva M, Jung F, Ortiz C, Lentini A, Csillag V, Graziano M, Nikolakopoulou I, Weglage M, Lazaridis I, Kim H, Lenzi I, Park H, Reinius B, Carlén M, Meletis K. Esr1+ hypothalamic-habenula neurons shape aversive states. Nat Neurosci. 2023 Jul;26(7):1245-1255.
Chen S, He L, Huang AJY, Boehringer R, Robert V, Wintzer ME, Polygalov D, Weitemier AZ, Tao Y, Gu M, Middleton SJ, Namiki K, Hama H, Therreau L, Chevaleyre V, Hioki H, Miyawaki A, Piskorowski RA, McHugh TJ. A hypothalamic novelty signal modulates hippocampal memory. Nature. 2020 Oct;586(7828):270-274.
Farrell JS, Lovett-Barron M, Klein PM, Sparks FT, Gschwind T, Ortiz AL, Ahanonu B, Bradbury S, Terada S, Oijala M, Hwaun E, Dudok B, Szabo G, Schnitzer MJ, Deisseroth K, Losonczy A, Soltesz I. Supramammillary regulation of locomotion and hippocampal activity. Science. 2021 Dec 17;374(6574):1492-1496.
Szőnyi A, Zichó K, Barth AM, Gönczi RT, Schlingloff D, Török B, Sipos E, Major A, Bardóczi Z, Sos KE, Gulyás AI, Varga V, Zelena D, Freund TF, Nyiri G. Median raphe controls acquisition of negative experience in the mouse. Science. 2019 Nov 29;366(6469):eaay8746.
The original impetus for this sleep essay was the idea that the basal ganglia could best be understood as a sleep and wake circuit [Kazmierczak and Nicola 2022]. After reviewing the rest of the brainstem sleep circuitry, it’s time to tackle the original problem.
Snr as a sleep/wake gate
Snr (substantia nigra pars reticulata) is the output node of the basal ganglia. It’s a set of GABA neurons that tonically suppress the majority of all brainstem motor areas including MLR (midbrain locomotor region), OT (optic tectum), and R.rs (hindbrain reticulospinal motor command) with corollary discharge to the thalamus. Snr can inhibit initiation of eating and motion [Rossi et al 2016], but don’t disrupt ongoing actions [Liu et al 2018]. Disruption of Snr can cause hyperactivity and insomnia [Geraschenko et al 2006]. The caudal Snr derives from hindbrain r1 (rhombomere r1 near the midbrain-hindbrain boundary) [Achim et al 2012], [Lahti et al 2015], [Partanen and Achim 2022], suggesting it may be evolutionarily old, possibly older than other basal ganglia regions.
Sleep as gating motive from action or sleep from action. Wake as disinhibiting sleep. Snr (substantia nigra pars reticulata).
As described in part 1 this essay, sleep suppresses senses, motivation and action. To implement this suppression, sleep could disconnect senses and motivation neurons from action neurons. In the above diagram, the gate is conceptual. The circuit could also inhibit the sense or action nodes directly instead of requiring specific gating neurons. This gating architecture has the advantage of simplicity because the sleep circuit can be localized in the gate, while the senses and actions can be mostly free of sleep circuitry.
As a preview, sleep neurotransmitters and peptides in BG (basal ganglia) include AD (adenosine), enk (enkephalin), MOR (μ-opioid receptor), and wake neurotransmitters include DA (dopamine), tac1 (tachykinin 1 aka neurokinin 1 aka substance P), dyn (dynorphin), and DOR (δ-opioid receptor).
If the vertebrate brain follows this architecture, Snr is well-placed to control that gate. Snr.m (medial Snr) projections have many collaterals to distinct motor areas and suppressing the wake-promoting areas covered earlier in this essay, which suggests widespread suppression as opposed to fine-grained control.
Snr.m gad2 connectivity. 60% of Snr.m inputs are from motor, motivation and wake areas. H.l (lateral habenula), H.stn (subthalamic nucleus), H.zi (zona incerta), M.pag (periaqueductal gray), OT.m (medial optic tectum), P.ge (external global pallidus), Ppt (pedunculopontine nucleus), R.rs (reticulospinal motor command), S.d (dorsal striatum), Snr.m (medial substantia nigra pars reticulata).
As the above diagram illustrates, despite its description as basal ganglia output, 60% of the gad2 (genetic marker), Snr.m inputs are outside of the basal ganglia, particularly from the midbrain (30%) and hypothalamus (10%) [Liu et al 2020]. Snr.m has two independent neuron types marked by gad2 and pv (parvalbumin), which are topographically organized with gad2 in Snr.m and pv in Snr.l (lateral Snr). While Snr.l.pv seems to be strictly motor related, Snr.m.gad2 are sleep related [Liu et al 2020]. However, [Lai et al 2021] reports Snr.l as sleep related.
Functional sleep and action requirements. Any ongoing action should suppress sleep, and sleep should suppress all actions.
Snr’s widespread motor and motivation connectivity suggests a possible primitive role in sleep. Sleep needs to suppress all actions, but any ongoing action needs to suppress sleep, because an animal shouldn’t fall asleep while eating or moving. It seems plausible that a primitive proto-vertebrate could have used Snr for sleep regulation without needing the rest of the basal ganglia.
Because astrocytes can integrate inputs spatially and temporally and are associated with sleep, it’s plausible that Snr astrocyte would be involved in this circuit. Interestingly Snr astrocytes are sensitive to dopamine and become hyperactive in the absence of dopamine [Bosson et al 2015] and are sensitive to glutamate from H.stn [Barat et al 2015].
Dopamine D2.i sleep / wake circuit
Although the independent Snr circuit is a functional sleep / wake gating circuit, it tonically inhibits the sense to action circuit, adding noise. An improvement to the circuit enables the gate when a signal is available, using the striatum to selectively open the gate. This circuit uses dopamine to open and close the gate. High dopamine is a wake signal and low dopamine is a sleep signal.
In the above diagram, Snr and S.d2 (D2.i associated striatum projection neurons) are sleep-promoting regions and S.d1 (D1.s associated striatum projection neurons) is a wake-promoting region. D2.i (inhibitory Gi-protein dopamine receptor) disconnects inputs, as opposed to inhibiting a neuron directly. When DA is available, S.d2 is disconnected, and S.d1 inhibits Snr, opening the gate. When DA is low, S.d2 is active, which inhibits S.d1, disinhibiting Snr, closing the gate and producing sleep. The D2i between S.d2 and S.d1 is from [Dobbs et al 2016].
The idea of the circuit is that the sense signal disinhibits itself during wake, but sleep prevents sense from disinhibiting itself. The minimal system only requires D2i circuits [Oishi et al 2017]. Wake enables the gate, and sleep disables the gate. Although I’ll cover D1s later, D2i is more fundamental because disabling D1s can be reversed by sufficient arousal, but disabling D2i can’t [Kazmierczak and Nicola 2022].
Note the diagram is somewhat incorrect, because direct S.d2 to S.d1 connection is weak [Tepper 2008]. Instead, S.d2 GABA inhibits S.d1 input at distal dendrites as opposed to inhibiting the neuron soma itself.
P.v ventral pallidum and S.core
While S.d2 neurons in model above suppresses motor for sleep, S.d2 in S.core (ventral striatum core aka nucleus accumbens) can produce sleep pressure by inhibiting the wake supporting P.v (ventral pallidum) [Oishi et al 2017]. P.v is a tonically active, wake-promoting nucleus, primarily inhibiting sleep areas or disinhibiting wake areas.
Sleep/wake control adding P.v as a tonic wake producing node. DA (dopamine), D2i (inhibitory Gi-coupled dopamine receptor), H.l (lateral hypothalamus), Hb.l (lateral habenula), M.pag (periaqueductal gray), Ppt (pedunculopontine nucleus – ACh), P.v (ventral pallidum), S.d1 (D1-associated striatum projection neuron), S.d2 (D2-associated striatum projection neuron), Snr (substantia nigra pars reticulata), V.mr (median raphe – serotonin), Vta (ventral tegmental area – dopamine).
P.v fill a similar wake-promoting role as S.d1, but unlike S.d1 it’s tonically active and affects the motivation loop of H.l, Hb.l, and Vta instead of gating sense from action. Where P.v supports general wake, S.d1 supports specific wake for an action. Like the previous basal ganglia sub-circuit, this sub-circuit only requires D2i receptors.
P.v promotes wake by inhibiting Hb.l sleep-producing system [Li et al 2023]. It also promotes wake through Vta by disinhibiting GABA interneurons [Li et al 2021]. (It could also disinhibit H.l orexin but I don’t have a reference).
In the model above, stimulating S.d2 inhibits wake-producing P.v, which disinhibits sleep-producing areas like Hb.l and inhibits wake-producing areas like H.l and Vta through GABA interneurons. Conversely, stimulating the D2i receptor by high DA inhibits S.d2, which disinhibits Pv, allowing it so promote wake. Disabling the D2i receptor activates S.d2, promoting sleep even with high dopamine [Qu et al 2010].
Note that S.d1 also connects to P.v and can produce wake [Zhang et al 2023]. P.v has multiple sub-populations with opposing functions. For example, it has both a hedonic hot spot for liked food and a cold spot for disliked food [Castro et al 2015]. For the sake of simplicity the diagram only shows a sleep-promoting path through S.d2, but there may be a wake-promoting path through S.d2 to an opposing P.v subpopulation.
D1s – stimulator dopamine receptors
Although using only D2i as a mode switch to the sleep path is functional, it can be improved by also enhancing the wake path with D1s (stimulatory Gs-coupled dopamine receptor).
D1s as enhancing the basal ganglia wake path. DA (dopamine), D1s (stimulatory Gs-coupled dopamine receptor), D2i (inhibitory Gi-coupled dopamine receptor), S.d1 (D1-associated striatum projection neuron), S.d2 (D2-associated striatum projection neuron), Snc (substantia nigra pars compacta – dopamine), Snr (substantia nigra pars reticulata).
The improved circuit works exactly like the D2i-only circuit but enhances the wake path when DA is available. Dopamine boosts both the signals from the sense to S.d1 and the signal from S.d1 to Snr [Salvatore 2024], [Kliem 2007], [Rice and Patel 2015]. When dopamine is available, it boots the sense to S.d1 signal with D1s, which more strongly disinhibits the gate by inhibiting Snr, which is also boosted by D1s.
The D1s in Snr and dopamine may be more important for motor suppression than dopamine in the striatum [Salvatore 2024]. In Parkinson’s disease and also normal aging, bradykinesia (slow movement) correlates with dopamine in Snr more closely than dopamine in the striatum. Motor symptoms in Parkinson’s disease don’t generally occur until striatal dopamine is reduced by 80%, but the effect on Snr is more immediate with only a small drop of dopamine.
Note that the Snc (substantia nigra pars compacta) to Snr dopamine comes from somatodendritic broadcast, not from an axon synapse. Snc dendrites in Snr produce dopamine to enhance the S.d1 to Snr connection.
Although the previous diagrams show the basic logic of the circuit, the basal ganglia use adenosine as a sleep-producing neurotransmitter, competing with dopamine.
Adenosine in striatum sleep
Adenosine is a product of the energy molecule ATP and is produced by neural activity, and also as a astrocyte transmission molecule. Although adenosine can accumulate in a circadian manner, particularly in P.bf (basal forebrain), it’s typically a shorter term sleep pressure. Caffeine is wake promoting by suppressing adenosine receptors.
Dopamine and adenosine are paired, opposing neurotransmitters in the basal ganglia: dopamine produces wake and adenosine promotes sleep. As an opposing signal to dopamine, the adenosine circuit is a flip version of the dopamine circuit.
Parallel adenosine sleep circuit in the basal ganglia. AD (adenosine), A1i (inhibitory Gi-coupled adenosine receptor), A2a.s (stimulatory Gs-coupled adenosine receptor), S.d1 (D1-associated striatum projection neuron), S.d2 (D2-associated striatum projection neuron), Snr (substantia nigra pars reticulata).
When adenosine is active in the above circuit, it cuts off S.d1 input and output and enhances S.d2’s suppression of S.d1. With S.d2 fully suppressed, Snr is free to suppress the gate and therefore suppress sleeping action.
Since adenosine is low in the morning, sleep is suppressed, which is enhanced by high ultradian morning dopamine. If A2a.s (stimulating Gs-coupled adenosine receptor) are stimulated in the striatum, the animal is more likely to sleep even in the morning [Yuan et al 2017], specifically in S.core not S.sh (ventral striatum shell aka nucleus accumbens) [Oishi et al 2017].
The dual signal system allows for interesting combinations at the boundary between sleep and wake. If adenosine is high with sleep pressing, then a large amount of dopamine motivation is required to continue wake. In fact, sleep deprivation down regulates D2i receptors, moving from the neuron membrane to the interior [Volkow et al 2012], which tips the balance toward sleep by diminishing the D2i-mediated wake signal. Caffeine inhibits both the A1i (inhibitory Gi-coupled adenosine receptor) and A2a.s receptors, tipping the balance to dopamine wake.
Dorsal striatum indirect path
The full S.d (dorsal striatum) path includes an indirect path, but this path may be more related to pure motor control, not sleep. As mentioned above, Snr divides into two populations Snr.l with pv neurons and Snr.m with gad2 neurons, and the Snr.l neurons are motor related, not sleep related [Liu et al 2020]. Similarly, the indirect path including P.ge (external globus pallidus) and H.stn (sub thalamic nucleus) may not be sleep related. Nevertheless, I’ll include it here, in case it is sleep related.
S.d model with indirect path included. DA (dopamine), D1s (stimulatory Gs-coupled dopamine receptor), D2i (inhibitory Gi-coupled dopamine receptor), H.stn (subthalamic nucleus), P.ge (external globus pallidus), S.d1 (D1-associated striatum projection neuron), S.d2 (D2-associated striatum projection neuron), Snc (substantia nigra pars compacta), Snr (substantia nigra pars reticulata).
Note that both P.ge and H.stn are tonically active, and they oscillate together at beta frequencies (roughly 10hz), which suppresses action. An excessive beta oscillation in this P.ge and H.stn circuit is a Parkinson’s disease symptom that suppresses motion and can also interrupt sleep. D2i receptors in H.stn mean that dopamine suppresses H.stn output [Shen et al 2012].
One significant experiment showed that lesioning P.ge increased wake by 40%, particularly eliminating normal circadian night-time sleep, replacing it with day-time like napping [Qiu et al 2016], which would suggest that P.ge is a major sleep center like Po.vl (ventrolateral preoptic area) [Vetrivelan et al 2010]. Note that this analysis would suggest that my basal ganglia sleep diagram is entirely wrong, because P.ge as a sleep center is basically incompatible with its position in the circuit.
P.ge – external globus pallidus
Lesioning P.ge increases wake by 40%, almost entirely eliminating circadian sleep [Qiu et al 2016]. However, this produces hyperactive chewing, weight loss, abnormal motor behavior and death in 3-4 weeks [Vetrivelan et al 2010]. Other manipulations of P.ge produce hyperactivity, abnormal movement, and odd stereotypical behavior [Gittis et al 2014]. So, it’s unclear to me that P.ge is a sleep center, but removing P.ge produces excessive action which then suppresses sleep.
In addition, P.ge is a heterogenous area with at least three major cell types with distinct projections and roles. Arkypallidal neurons project strongly and exclusively to the striatum. Lhx6 neurons project strongly to Snc and to some areas of H.stn, excluding the center. Pv neurons project to all of H.stn and also to T.pf (parafascical thalamus) [Gittis et al 2014].
Distinct projection neuron types of P.ge. H.stn (subthalamic nucleus), P.ge (external globus pallidus), Snc (substantia nigra pars compacta), Snr (substantia nigra pars reticulata), Spn (striatal projection neuron), Spv (pv marked striatum interneuron), T.pf (parafascicular thalamus).
With three projection types, it’s possible that they have entirely separate functions. For example, the lhx6 projections are functionally compatible with a sleep promoting role, and lhx6 neurons in H.zi (zona incerta) are sleep promoting [Liu et al 2017].
Liu K, Kim J, Kim DW, Zhang YS, Bao H, Denaxa M, Lim SA, Kim E, Liu C, Wickersham IR, Pachnis V, Hattar S, Song J, Brown SP, Blackshaw S. Lhx6-positive GABA-releasing neurons of the zona incerta promote sleep. Nature. 2017 Aug 31;548(7669):582-587. doi: 10.1038/nature23663. Epub 2017 Aug 23.
The first two parts of this essay were a general overview of the necessity of sleep and some of the properties. Here I’m going over some of the brainstem circuits that control sleep.
Wake ignition
Waking requires intrinsic motivation because sleeping places the animal away from distraction, to an extreme in hibernation. A short nap, as is more typical in the waking period, needs to end without needing external stimulus or an internal one like hunger. What’s needed is an internal ignition source to drive wake and motivation.
In rodents, if the area around R.pb (parabrachial nucleus in r1) is lesioned, the animal remains in a coma [Fuller et al 2011]. For humans, a study of coma showed a pattern of the same area as consistently being destroyed [Grady et al 2022]. However, the exact cells aren’t known, and other studies that lesion R.pb for conditioned taste studies don’t produce coma. Still, this site seems a likely ignition area.
Possible wake ignition subcircuit. R.pb is the main ignition source and wakes motivational areas like H.l. H.l (lateral hypothalamus), N5 (trigeminal nerve), N10 (vagus nerve), Nsp (spinal cord), R.pb (parabrachial nucleus), R.pz (parafacial zone).
The above diagram shows a possible wake ignition circuit. The area around R.pb is the main wake ignition node. R.pb produces wake by stimulating motivational areas like H.l (and others).
It’s not known if the R.pb area is self-igniting or if astrocytes in the area are critical, or if it uses peripheral wake signals such as N5 (trigeminal nerve), N10 (vagus nerve), or N.sp (spinal cord or other periphery) [Grady et al 2022]. In the fruit fly Drosophila specialized peripheral leg neurons can promote daytime sleep [Jones et al 2023]. These specialized neurons are distinct from sensor or motor neurons. In addition peripheral neurons from Drosophila PPM area are wake promoting [Satterfield et al 2022]. So, it seems plausible that peripheral nerves such as N5, N10, or N.sp could have similar wake-producing neurons, although this is entirely speculative.
If R.pb is a wake-ignition system, then sleep needs to suppress it, whether by internal clock regulation, or external suppression. In the hindbrain, R.pz (parafacial zone near N7 and r5 / r6) suppresses R.pb to create sleep [Anaclet et al 2014]. Disabling R.pz decreases NREM sleep by 30% [Erikson et al 2019].
Hindbrain (rhombomere) sleep
R.rs (reticulospinal) neurons in the caudal hindbrain (medulla, r6-r8) have both wake and sleep effects. Because R.rs are motor control neurons, they need to suppress sleep while they’re active, but the same area also contains sleep promoting areas. So, when experimenters stimulate the area, the animal remains awake, but immediately following the end of stimulation the animal sleeps, because the sleep-inhibition is removed. [Teng et al 2022]. These R.rs neurons (ventrolateral medulla) send collaterals to Po.vl (ventrolateral preoptic area), which is a forebrain sleep / wake area.
Midbrain sleep
A specific nucleus near N3 (oculomotor nerve) and associated motor areas (Edinger-Westphal) is a sleep promoting area. Stimulating it increases NREM [Zhang et al 2019]. (I’m just noting this for reference. I don’t understand how this area connects with other sleep areas.)
Ventral preoptic area
Although wake-maintaining areas are widely distributed, and much of sleep-circuitry is postponing sleep for ongoing actions, sleep-promoting area are more rare. One sleep-promoting area is Po.vl (ventrolateral preoptic area) and Po.mn (median preoptic area), which are adjacent area. Po.vl is inhibitory GABA and inhibits neurotransmitter wake areas like V.lc (locus coeruleous – norepinephrine), Vta (dopamine), V.dr (serotonin), M.pag.v (which is V.dr but refers to a dopamine area), and Ppt (pedunculopontine nucleus – acetylcholine) and P.ldt (laterodorsal tegmental nucleus – also acetylcholine).
Po.vl sleep-promoting area suppresses wake-promoting areas. H.l (lateral hypothalamus), Po.vl (ventrolateral preoptic area), R.pb (parabrachial nucleus), R.pz (parafacial zone), V (wake-neurotransmitter areas including dopamine, histamine, serotonin, acetylcholine, orexin).
In the diagram above, Po.vl promotes sleep by inhibiting wake-promoting areas, here represented by H.l and V, where V includes the neurotransmitter wake areas. This Po.vl function is one pole of the flip-flop analogy [Saper et al 2001], driving sleep transitions faster and supporting continuous sleep, avoiding fragmentation.
Lateral Hypothalamus
H.l is a sleep wake hub [Gazea 2021] with both wake-promoting peptide orexin, and GABA neurons promoting wake. The orexin peptide is wake-related, because if it’s missing, people and animals develop narcolepsy and cataplexy. H.l orexin neurons project to other wake-promoting areas like V.lc, Ppt, V.dr, Vta, and is believed to sustain wakefulness.
Stimulating orexin neurons does produce wake after sleep, but only after 10-20 seconds, so these aren’t directly wake producing, but more wake facilitating. In contrast, V.lc norepinephrine neurons produce wake in 2 seconds [Yamaguchi et al 2018].
In addition, a study suggested that human orexin is low at dawn, a time when people are active and twos to peak at dusk [Mogavero et al 2023]. So orexin’s role is something more complicated than simply a wake-promoting peptide. (Note: this study seemed somewhat unreliable. I’d like to see a more detailed orexin over time study for rodents, where measurements can be more precise).
Other sleep and wake neurons exist in H.l without the orexin [Heiss et al 2018]. For example, some Vta GABA neurons that express SST (somatostatin) store sleep requirements for up to 5 hours, and signal the extra sleep need to H.l [Yu et al 2019].
Lateral hypothalamus and value neurotransmitters as a wake hub. Hb (habenula), H.l (lateral hypothalamus), pineal (pineal gland), R.pb (parabrachial nucleus), V (wake-promoting neuropeptides)
The above diagram shows two complementary roles for H.l. First, H.l can suppress Hb.l circadian sleep-promoting path with H.l orexin projections to Hb that inhibit anaesthesia [Zhou et al 2023] and promote aggressive arousal [Flanigan et al 2020].
The positive feedback loop from H.l to Hb (habenula) to V and back to H.l sustains wake. The gain for positive feedback could vary in circadian cycles. A high gain in the morning could produce full wake even with little activity. A low gain at night would make it harder to sustain wake.
Misc notes: Sleep preparation is a distinct, complicated pre-sleep behavior. One trigger seems to be from F.pl (pre limbic frontal cortex) SST neurons to H.l [Tossell et al 2023]. Astrocytes also seem to be involved with H.l wake, are active when waking and promoting wakefulness [Cai et al 2022]. H.pv is also sleep promoting and if it’s knocked out, significant daytime sleep increases, particularly in the morning [Chen et al 2021]. In contrast, the posterior hypothalamus has astrocytes that increase sleep at night [Pelluru et al 2016].
Vta sleep/wake glutamate and GABA
For the moment, let’s ignore Vta (ventral tegmental area) dopamine. Vta includes glutamate and GABA neurons that derive from r1 (hindbrain rhombomere 1) [Lahti et al 2016] that enhance wake with glutamate [Yu et al 2019] and enhance sleep with GABA [Chowdury et al 2019]. The tail of Vta, RMTg (rostromedial tegmental are in r2 / r3) is essential for NREM sleep [Yang et al 2018].
Vta glutamate, GABA, and DA all control sleep using H.l and S.msh (medial shell of the ventral stratum aka nucleus accumbens). As noted above, H.l is a coordinator of sleep and wake with not only orexin but also GABA and glutamate. Inhibiting Vta GABA bypasses sleep homeostasis, producing a mania state during circadian wake times [Yu et al 2021], [Yu et al 2022].
Vta glutamate stimulation promotes continuous wake independent of DA [Yu et al 2019], via projections to H.l, S.sh, and P.v (ventral pallidum), particularly the NOS1 cells.
Vta.g (GABA neurons of Vta) stimulation encourages NREM sleep. If Vta.g are inhibited, the animal remains 100% awake for hours, during the normal wake period, not the normal sleep period [Yu et al 2022]. The Hb.l projection to Vta.g is required for the anesthetic propofol to work [Gelegen et al 2018].
Interestingly, a specific SST (somatostatin) subset of Vta GABA retains a future sleep requirement from social defeat, where social defeat produces extra sleep. When the rodent loses a conflict, these Vta SST neurons have elevated calcium for up to five hours, and then the animal sleeps, these neurons fire to H.l, extending sleep duration [Yu et al 2019]. Speculating here, this multi-hour memory suggests a possible astrocyte involvement.
Returning the dopamine. Vta dopamine produces wake, while Vta dopamine inhibition produces sleep with nesting behavior [Eban-Rothschild et al 2016], as opposed to immediate collapse like narcolepsy. Low dopamine in a behaviorist experiment produces long decays, difficulty in locomotion and sleepiness [Nicola 2007]. However, other studies argue that dopamine itself is not wake promoting [Takata et al 2018].
Habenula
As mentioned in a previous post, Hb (habenula) is a sleep-promoting area as a motor-inhibiting area driven by the pineal gland and extending melatonin’s role [Hikosaka 2012]. This sleep promoting area is in a positive feedback loop with the wake-promoting neurotransmitters and peptides.
Pineal gland through habenula as promoting sleep by suppressing motivation and motor action. Hb (habenula), V (wake-promoting neurotransmitters and peptides).
This above diagram is a different perspective on the prior H.l diagram, where I’ve merged H.l into V and made the motor and motivation suppression explicit. Here the wake-promoting neurotransmitters gate motivation and motor, extending the role of melatonin, which suppresses action.
Active actions promote wake and suppress sleep, like the R.rs wake efferent copies [Teng et al 2022], by stimulating the value neurons. In turn, the value neurons suppress the habenula such as Vta to Hb.l [Webster et al 2021] and serotonin inhibiting Hb.l [Tchenio 2016], H.l orexin and GABA also inhibit Hb.l [Flanigan 2020], [Gazea 2021]. As mentioned above, these positive feedback loops sustain wake despite sleep pressure.
Summary circuit
Putting these components of the sleep/wake circuit together produces something like the following, where I’ve emphasized the habenula to show how that subsystem fit into the whole circuit.
Sleep/wake circuit emphasizing the pineal, melatonin habenula path. Hb (habenula), H.l (lateral hypothalamus), S/P (basal ganglia), Po.v (ventrolateral and median preoptic areas), R.pb (parabrachial nucleus), R.pz (parafacial zone).
As before, the circadian sleep drive from the pineal gland drives the habenula, which inhibits wake neurotransmitters, which inhibits motivation and action using the basal ganglia as a gate. Ongoing action sustains wake against habenula-driven sleep pressure.
The full diagram includes the wake-ignition circuit from R.pb, the sleep-sustain circuit in Po.v (Po.vl and Po.mn), and the wake-sustain circuit in H.l and V. As a reminder, this model is highly simplified and really only serves as a skeleton to organize the various brainstem sleep systems.
Cortical slow wave sleep
Although I’m trying to avoid the cortex as long as possible, studies use cortical slow waves as a sleep marker, so it’s inescapable. Cortical slow waves are globally synchronized neuron firing between around 0.5Hz to 4Hz. The slow wave firing has no information content, but the oscillations may help clear the cortex of metabolic toxins.
During wake, neurons expend to fill the intercellular space because of the neuron’s ion gradients. Filling the intercellular space means the CSF (cerebral-spinal fluid) can’t clear metabolic toxins [Xie et al 2013]. Slow wave sleep shrinks the neurons allowing fluid to fill the intercellular space, and the oscillations may even help with fluid circulation [Fultz et al 2019].
Cortical sleep appears strongly coupled to astrocytes. Astrocyte calcium precedes slow waves in the cortex [Poskanzer and Yuste 2016]. Astrocytes may even organized SWS waves across the cortex, using electrical gap junctions to connect to astrocyte neighbors [Vaidyanathan et al 2021].
Wake signals driving cortical wake. C (cortex), H.l (lateral hypothalamus), Hb (habenula), P.bf (basal forebrain), V (wake neuropeptides)
The above diagram shows a simplified cortical wake circuit, although the cortex is also affected by wake neurotransmitters norepinephrine, serotonin and dopamine. In this model the cortex is mostly an appendage of the brainstem sleep circuit, waking when the brainstem wakes.
P.bf (basal forebrain) is a set of GABA and ACh nuclei that activate the cortex, hippocampus, and olfactory bulb. Although P.bf is identified by its ACh neurons, the GABA projections seem to be more important for wake.
Notes: Local cortical sleep pressure is signaled with GABA [Alfonsa et al 2023]. Parts of the cortex can sleep independently [Krueger et al 2013].
Ppt and P.ldt ACh and wake
Although I’ve lumped Ppt (pedunculopontine nucleus) and P.ldt (laterodorsal tegmental nucleus) with the “V” wake promoting areas, they deserve a special mention because of their connection and similarity with P.bf. Ppt and P.ldt are ACh ganglia near the isthmus midbrain-hindbrain boundary. Ppt is part of the MLR, showing the tight connection between locomotion and wake. Ppt feeds into P.bf, the striatum, and other locomotive regions like H.stn.
Interestingly, all Ppt neurons self-generate gamma oscillations through intrinsic channels [Garcia-Rill et al 2015]. So it could be an ignition source of gamma activation in the striatum and cortex.
Yu X, Li W, Ma Y, Tossell K, Harris JJ, Harding EC, Ba W, Miracca G, Wang D, Li L, Guo J, Chen M, Li Y, Yustos R, Vyssotski AL, Burdakov D, Yang Q, Dong H, Franks NP, Wisden W. GABA and glutamate neurons in the VTA regulate sleep and wakefulness. Nat Neurosci. 2019 Jan;22(1):106-119.
As mentioned in the previous post, sleep is often divided into circadian sleep and homeostatic sleep, although this model is an oversimplification in part because of the metabolic cycles [Borbély et al 2016]. Despite the caveats, I think starting from circadian circuits is a good start.
Also as mentioned previously, circadian cycles may have started as an oxidation-reduction cycle to project from oxygen’s toxicity after the Great Oxidation Event [Edgar et al 2012]. One of the early solutions is melatonin, a powerful natural antioxidant [Tosches et al 2014].
Melatonin
Melatonin exists in almost all animals except sponges. Along with its antioxidant properties, it signals for the zooplankton diel vertical migration, swimming toward the light at dusk and sinking at night [Tosches et al 2014].
System for melatonin-controlled zooplankton vertical migration. ACh (acetylcholine).
In the migrating zooplankton, melatonin triggers ACh (acetylcholine) neurons, which rhythmically spike and these spikes disrupt the cilia, disorganizing them and allowing the plankton to sink.
Reptile and mammal complications
As a complication to understanding the vertebrate circuits, both reptiles and mammals have sleep requirements at odds with aquatic vertebrates. Because land temperatures change more than water temperatures, and reptiles are cold-blooded, their sleep and wake is necessarily strongly tied to temperature as well as the common light/dark connection [Rial et al 2022]. So, sleep and temperature are highly correlated, which makes the Poa (preoptic area) combination of temperature and sleep functions more reasonable than a seemingly random combination.
Mammals have the additional complication of the evolutionary nocturnal bottleneck [Rial et al 2022], meaning the simple heuristic of nighttime melatonin for sleep isn’t sufficient. The pineal melatonin is still at night for nocturnal animals [cite], and the light signaling needs to flip. Although diurnal mammals are no longer nocturnal, their clock circuitry retains the heritage of a nocturnal flip.
As a specific example, all non-mammalian vertebrates use the pineal gland as a circadian oscillator, not H.scn (subthalamic nucleus) [Vatine et al 2011].
Pineal gland and habenula
The pineal gland in the midbrain is the vertebrate’s main source of melatonin. Evolutionarily, the pineal gland is derived from photosensitive cells that directly convert light and dark into melatonin. In non-mammals, the pineal gland is still photosensitive. In the zebrafish, the pineal photoreceptor is still effect and entrains circadian cycles [Vatine et al 2011], and an analogous region in the non-vertebrate chordate Amphioxus provides a similar function, showing the pineal gland’s conserved function in vertebrates [Lacalli 2022].
Hb.m and Hb.l (medial and lateral habenula) derive from the pineal complex, and may have originally been effectors of the pineal gland, serving a nervous function analogous to melatonin [Hikosaka 2010]. Hb.l in particular is well-suited to control neurotransmitters associated with wake, such as dopamine from Vta (ventral tegmental area), serotonin from V.dr and V.mr (dorsal and medial raphe), and norepinephrine from V.lc (locus coeruleus). Note that melatoninAs explored in essay 20, Hb.m is involved in primitive phototaxis and chemotaxis and is well-placed to inhibit those actions during sleep.
Habenula control of sleep by gating motive from action. Hb (habenula), pineal (pineal gland).
In the above diagram, a primitive habenula function is to suppress sensation, motivation and action for sleep by suppression wake-supporting neurotransmitters. Although the diagram illustrates the habenula as disconnecting motive from action, it could also disconnect sense from action, as in phototaxis or chemotaxis in Hb.m.
Hb.m includes an internal entrainable circadian clock, unlike Hb.l. The Hb.m clock is necessary for ultradian foraging. The foraging ultradian is around four hours, generally on waking. Both dopamine and NE (norepinephrine) are elevated [Wang et al 2023] and reciprocally the circadian clock is set by dopamine and NE [Salaberry and Mendoza 2022].
Some misc notes: Hb.l is required for some anesthesia (propofol) and stimulating Hb.l strongly induces NREM, and suppresses motor [Gelengen et al 2018]. Hb.l stimulus produces NREM [Goldstein 1983]. Hb.l is more active mid and late day and early night [Aizawa et al 2013] (possibly producing morning ultradian activity). Hb.l manipulation produces wake fragmentation in the wake period and sleep fragmentation in the sleep period via orexin in H.l [Gelengen et al 2018].
Cell clocks
As mentioned in the introduction, the oxidation-reduction protection may have led to the development of cellular clocks. Essentially all cells have circadian cycle in protein expression, including metabolic and detoxification cells in the liver, heart, kidneys and digestion [Dibner et al 2010], even including gut bioflora. The clocks are synchronized by multiple signals, including feeding patterns, but most studied by light.
For example, dopamine is under clock control and is modulated by melatonin [Ashton and Jagannath 2020]. In S.v (ventrial striatum aka nucleus accumbens) dopamine is at a daily low at night. DAT (dopamine transporter), affected by cocaine, is regulated by clock genes [Alsonso et al 2021], possibly under control of astrocytes. Dopamine is particularly tonically high in early morning before eating with an ultradian cycle of about four hours. Two four-ish hour dopamine cycles are known: the FEO (food entrainable oscillator), which produces pre-feeding activity [Dibner et al 2010], and MASCO (methamphetamine-sensitive circadian oscillatory) [Tataroglu et al 2006], which may be the same system.
The retina itself is under circadian control, modulated by dopamine and D2i (inhibitory Gi-coupled dopamine receptor) [Yujinovsky et al 2006], including in frogs [Cahill and Besharse 1991].
And astrocytes in S.v are under circadian cell clock control [Becker-Krail et al 2022]. Astrocytes are well-placed to manage sleep because they have widespread connections to many synapses and are connected to other astrocytes with gap junctions, allowing for integration over time and space and widespread broadcast signaling.
H.scn circadian entrainment
The circadian system has three distinct components that can either work on their own or work together:
Cell clocks
Light / dark photoreceptors or feeding signals and behavior
Entraining the cell clock to the signal (zeitgeber)
If the eye area of the mollusk sea hare is lesioned, circadian entrainment is eliminated, but because of other photoreceptors, the animal still follows light and dark cycles as long as the light changes. The deficit is only exposed when the lesioned mollusks are placed under continual dark [Vorster et al 2014], [Newcomb et al 2014]. Similarly, in zebrafish many cells are photoreceptive without entraining the cellular clocks.
Mammals use H.scn (suprachiasmatic nucleus) to coordinate circadian cellular clocks. The H.scn name is important, but it’s located above the optic crossing (suprachiasmatic) and developmentally the retina develops from the hypothalamus adjacent to H.scn.
Abstract representation of the mammalian brain highlighting the proximity of the retina and H.scn. arc (H.arc – arcuate nucleus), C (cortex), CB (cerebellum), H.l (lateral hypothalamus), ip (R.ip interpeduncular nucleus), mb (H.mb mammillary bodies), MHB (midbrain-hindbrain boundary), P (pallidum), S (striatum), scn (suprachiasmatic nucleus), sum (supramammillary nucleus), Vta (ventral tegmental area), r1 (rhombomere 1), ZLI (zona-limitans intrathalamica)
The diagram above shows the rough location of the retina development area and H.scn, which both develop from the hypothalamus. A primitive eye with only a few photoreceptors would have been part of the hypothalamus, and like the mollusk the photoreceptor would be near the clock entrainment circuit that became H.scn.
The H.scn clock signal is somewhat indirect, with an interim projection to H.scz to H.dm (dorsomedial hypothalamus) and finally to H.l (lateral hypothalamus) for wake and Po.vl (ventrolateral preoptic area) for sleep. H.scn uses dopamine from Vta as part of its synchronization [Grippo et al 2017].
Ultradian DA – morning foraging
The sleep / wake cycle has an additional boost during normal foraging times such as immediately after waking. In the subjective morning (dark for rodents), wake is encouraged, homeostatic sleep is suppressed, and dopamine levels are higher. After the foraging boost ends, but still in the wake period, tonic dopamine levels drop and the animals take more frequent naps. This hut radian boost of about four hours affects learning and behavior as well as modulating drug abuse [Ruby et al 2013].
Sleep / wake cycle showing morning boost. ZT (zeitgeber time).
Because this ultradian foraging boots wake and suppresses sleep significantly, studies that stimulate or inhibit sleep and wake can specifically affect the ultradian boost without affecting other sleep / wake periods. So it’s very important to look at the hourly effects because the experimental modulation might reduce the foraging boost specifically, but a summary might show a general sleep increase.
H.scn circadian entrainment uses dopamine. DA from either Vta [Grippo et al 2017], [Tang et al 2022] and/or H.sum (supramammillary nucleus) [Luo et al 2018] can entrain food circadian cycles. Note that since the dopamine “A10” area extends beyond the Vta to include H.sum and M.pag.v on opposite ends of the Vta, these studies may be reporting the same area.
The inhibitory neurotransmitter GABA is associated with sleep, and many sleep drugs are GABA stimulants. GABA neurons in Snr (substantia nigra pars reticulata), H.zi (zona incerta), Vta, and Po.vl are all associated with sleep. As mentioned above GABA from mitochondria and in Hydra are used as a sleep promoting neurotransmitter.
While GABA is associated with sleep, other major neurotransmitters like NE, DA, 5HT (serotonin), ACh (acetylcholine) and histamine are associated with wake maintenance of the execution of wakefulness. As discussed in the previous post, ongoing actions need to suppress sleep. NE, DA, and 5HT are all maintain wake while the animal is active and drop when the animal is winding down activity to sleep. Cortical wake requires activity in ACh-rich area in Ppt (pedunculopontine nucleus), P.ldt (laterodorsal tegmental nucleus), and P.bf (basal forebrain).
Produced by H.l, orexin (aka hypocretin) appears to be a wake-maintenance peptide since removal of orexin produces narcolepsy. H.l orexin projects to essentially all of the other wake-maintaining neurotransmitters, including NE, DA, 5HT and ACh. Orexin is slow, waking after tens of seconds, while stimulating V.lc NE is around two seconds [Yamaguchi et al 2018]. On counterargument is that orexin can ramp later in the day [Grady et al 2006], [Mogavero et al 2023], which would suggest that it’s not part of the ultradian foraging system, although it’s also highly tied to foraging. (Suggesting I need to read more articles to see if the contradiction has been resolved.)
Although orexin is the most dramatic of H.l wake, H.l also includes wake and sleep producing GABA and glutamate neurons that may be even more important for wake, independent of the orexin function. Unfortunately, H.l is complex enough that the different functions haven’t been fully pulled apart.
Adenosine is a sleep-promoting molecule derived from the energy molecule ATP, and has extensive receptor throughout the brain, notable in the striatum. Because it’s a product of ATP, it measures local neural activity and possibly sleep need. Its measurement of global brain activity for homeostatic sleep seems more questionable, but adenosine does accumulate throughout the wake period in P.bf [Porkka-Heiskanan et al 2000].
Inflammation peptides like IL-1β are also sleep-promoting [Imeri and Opp 2009]. In addition to their inflammation-related sleep, they seem to be part of normal homeostatic sleep signaling. In Drosophila sleep-need astrocytes produce IL-1β as a signaling peptide [Blum et al 2021]. In zebrafish, sleep deprivation correlates with immune signaling [Williams et al 2007].
Next: ignition and maintenance circuits
After this general discussion on sleep wake, the next post will cover some of the specific sleep and wake circuits, particularly those associated with wake ignition, wake maintenance and sleep maintenance.
Edgar RS, Green EW, Zhao Y, van Ooijen G, Olmedo M, Qin X, Xu Y, Pan M, Valekunja UK, Feeney KA, Maywood ES, Hastings MH, Baliga NS, Merrow M, Millar AJ, Johnson CH, Kyriacou CP, O’Neill JS, Reddy AB. Peroxiredoxins are conserved markers of circadian rhythms. Nature. 2012 May 16;485(7399):459-64.
Rial RV, Canellas F, Akaârir M, Rubiño JA, Barceló P, Martín A, Gamundí A, Nicolau MC. The Birth of the Mammalian Sleep. Biology (Basel). 2022 May 11;11(5):734. doi: 10.3390/biology11050734.
Because sleep is a global state that suppresses senses and actions, its control circuitry affects essentially all neural systems. For example, an article on dopamine and S.v (ventral striatum aka nucleus accumbens) suggested that dopamine acts more like a wake signal than an abstract reward signal [Kazmierczak and Nicola 2022]. If that explanation is accurate, then understanding the sleep system is a prerequisite for understanding the whole system.
From the study, low dopamine caused the rodents to either fall asleep normally or collapse in cataplexy, depending on whether D1s (stimulating Gs-coupled dopamine receptor) or D2i (inhibitory Gi-coupled dopamine receptor) were disabled. Many studies omit qualitative behavior like animals falling asleep, reporting only statistical summaries of success or failure.
The Great Oxidation Event
Sleep exists for essentially all animals including primitive animals like hydra and even single celled eukaryotes. Beyond sleep, oxidation-reduction cycles exist even for bacteria. 2.5 billion years ago in the Great Oxidation Event when photosynthesis created the toxin oxygen, most life died except for like that developed defenses against oxidation and ROS (reductive oxygen species). One of these cellular defenses was an oxidation-reduction cycle to spend time repairing oxygen damage. Cellular clocks developed around these primitive, conserved oxidation-reduction cycles [Edgar et al 2012].
Mitochondria in eukaryotes produce additional toxic oxygen ROS. One general sleep theory proposes that mitochondria force sleep on their hosts to allow for repair [Hartman and Kempf 2023]. In essentially all cells the cell clock and the mitochondrial clock are in sync [Scrima et al 2016]. In this model, sleep repairs oxidation damage in a quiet, low energy mode. Mitochondria produce GABA to signal to the host cell for its sleep need [Adams and O’Brien 2023]. GABA is the main inhibitory neurotransmitter, possibly directly inhibiting the containing neuron.
In the cortex a more sophisticated system passes damaged mitochondria from neurons to astrocytes, when then modulate sleep [Haydon 2017]. Astrocytes strong coupling between sleep and neural activity is important in many brain areas. In particular, astrocytes emit sleep transmitters adenosine and GABA, and connect to neighboring astrocytes with gap junctions to integrate sleep pressure spatially and temporally. Astrocyte can emit adenosine and GABA, both sleep signals. So, sleep can’t be treated as a straight neural circuit without considering the actions of astrocytes.
Beyond the brain, metabolic cells such as the liver and even gut microflora have circadian cycles and these metabolic cycles work best when synchronized with sleep [Borbély et al 2016].
Sleep basics
While sleep in mammals can be detected by slow waves in the cortex, a more general criteria is necessary to cover insects like Drosophila and worms like C. elegans. The following properties are generally used to identify sleep:
Behavioral quiescence
Sensory inhibition
Sleep position
From an implementation perspective, sleep has a global coordination problem because all processes need to sleep simultaneously. In contrast, waking processes such as foraging only needs to activate task-relevant areas, and other areas can rest outside of a general sleep state. Columns in the cortex, for example, can fall into a slow wave state while the animal is awake. Although no lesion of the brain produces a wake-only state [Krueger et al 2013], so there is no single sleep center, sleep requires global coordination.
Inhibit the link from stimulus to response
Inhibit intrinsic motivation
Inhibit cognitive processes
Two process model of sleep
The two process model of sleep considers circadian and homeostatic as two separate processes driving sleep. In addition, the animal’s activity can postpone sleep [Yamagata et al 2021]. Circadian sleep handles the major daily sleep need while homeostatic sleep covers local sleep needs.
Criticisms of this model point out metabolic anabolic and catabolic cycles are more related to feeding cycles than light cycles [Borbély et al 2016]. The presence of cell clocks in most cells suggests that circadian isn’t a global requirement. In addition, ultradian (4h) feeding cycles cause food anticipatory activity [Dibner et al 2010].
As a specific counterexample, the snail sleep can be model well by simple stochastic oscillator between wake and quiescence [Stephenson 2011]. In contrast zooplankton follow a clear circadian migration between light and dark [Tosches et al 2014].
From a circuit perspective, the two process model has value because some areas like Hb (habenula), Po.vl (ventrolateral preoptic area), and H.scn (suprachiasmatic nucleus) are more easily understandable from a circadian perspective.
Bistable sleep and wake
Although it might sound obvious that sleep and wake are distinct states, implementing this bistable system requires coordination. Violations of this bistability are unusual, like sleepwalking. As in essay 27, where the state transition between seeking food and eating required a circuit using H.stn (subthalamic nucleus) and Snr (substantia nigra pars reticulata), the sleep and wake circuits need circuits to manage their distinction and transition.
The sleep/wake transition needs to have high gain to avoid metastability.
Clear state transitions try to avoid metastability, a transitional non-state between the target states. Metastability always exists, but can be minimized by increasing the feedback gain between the states. A high gain, tight transition minimizes the probability of a metastable state. In the mammalian brain, positive feedback and lateral inhibition in Po.vl (ventrolateral preoptic area) and H.l.ox (orexin area of lateral hypothalamus) help make the switch tighter. [Saper et al 2001] calls this a flip-flop with the similarity to bistable electrical latches, where a high gain to avoid metastability is also very important to maintain binary values with a continuous voltage.
While high gain and lateral inhibition is important for sleep, an additional concept called hysteresis is also important to create long continuous sleep bouts and avoid sleep fragmentation.
Hysteresis: sticky switches
In a naive implementation of homeostatic sleep, the animal sleeps when sleep pressure rises past a threshold, and wakes when the pressure drops. Unfortunately, this system could quickly oscillate, where a short nap of a few seconds crosses the threshold and wakes the animal, which quickly tires and takes a new nap. To avoid this fragmented sleep, the threshold needs to be sticky: it’s harder to wake when the animal sleeps, and harder to sleep when the animal wakes.
Hysteresis for sleep pressure.
This kind of sticky switch is called hysteresis. The threshold for switching states depends on the current state.
Since sleep inhibits sensory input, noises that would keep an animal awake are ignored. Since sleep inhibits actions, the animal is unlikely to run into a situation that requires action. On the other side, any ongoing action will maintain wake. The transition to sleep needs to be slow to ensure that all actions have completed. In addition, long lasting peptides like orexin from H.l can maintain wake for minutes, ensuring a minimum wake bout length.
Note that circadian sleep process is another solution to the sleep oscillation problem. Because time is inexorable, circadian sleep time also shifts the sleep threshold, making it increasingly difficult to sustain wake.
Sleep and wake asymmetry
Sleep and wake are asymmetrical, unlike a symmetrical flip-flop. Any ongoing action needs to maintain wake, and falling asleep is a slow, decaying process, but waking needs to be fast when responding to an alarm. This asymmetrical slow drop to sleep and quick rise to wake is reflected in neurotransmitter levels like dopamine [Zhang et al 2023].
In terms of circuitry, an area around R.pb (parabrachial nucleus) is required for wake. If that area is lesioned, the animal remains in a coma [Fuller et al 2011]. There is no equivalent sleep area that produces a wake-only state when lesioned [Krueger et al 2013].
Next: circadian
After the general discussion of sleep, I think exploring the circadian aspect of sleep is a good direct. Circadian sleep is an ancient system, existing in zooplankton and preexisting more complicated sleep systems.
Edgar RS, Green EW, Zhao Y, van Ooijen G, Olmedo M, Qin X, Xu Y, Pan M, Valekunja UK, Feeney KA, Maywood ES, Hastings MH, Baliga NS, Merrow M, Millar AJ, Johnson CH, Kyriacou CP, O’Neill JS, Reddy AB. Peroxiredoxins are conserved markers of circadian rhythms. Nature. 2012 May 16;485(7399):459-64.
Yamagata T, Kahn MC, Prius-Mengual J, Meijer E, Šabanović M, Guillaumin MCC, van der Vinne V, Huang YG, McKillop LE, Jagannath A, Peirson SN, Mann EO, Foster RG, Vyazovskiy VV. The hypothalamic link between arousal and sleep homeostasis in mice. Proc Natl Acad Sci U S A. 2021 Dec 21;118(51):e2101580118.
Although the touch sense is a baseline implementation of obstacle avoidance, the essay’s animal should avoid obstacles before bumping into them, but it needs a new, ranged sense. The aquatic vertebrate lateral line senses obstacles, peer, predators, and prey at a distance of around the animal’s length. The lateral line is a basal vertebrate sense, lost in land vertebrates that’s a twin sense to weak electro sensation. Adding a lateral line sense to the toy model will let the simulated animal avoid walls before bumping into them.
Lateral line sense
The lateral line is an aquatic-only primitive vertebrate sense lost in terrestrial animals. Like mouse whisker, inner ear hearing and vestibular sensing, it’s a hair-based object-distance sensor that senses obstacles, peers, predators, and prey with a distance of about the animal’s length. Unlike mouse whiskers but like hearing and vestibular, the lateral line doesn’t touch the obstacle directly, but senses water movement, and from that motion can infer the presence of obstacles. The lateral line sensors are hair-like neuromasts that line the animal’s body along the entire length [Braun and Coombs 2000], [Chagnaud et al 2017].
Illustration of the lateral line sense from the animal (blue) sensing a nearby obstacle (grey wall at right).
Although the diagram above implies that the lateral line sense is a simple distance measurement, the actual sense is a complicated measurement of fluid dynamics and how water flow changes near obstacles. To further complicate the sense, the animal’s own motion changes the water flow. All vertebrates with the lateral line have a cerebellum-like nucleus to cancel out self motion called MON (medial octavolateral nucleus). Electrosensing fish have another cerebellum-like nucleus to cancel its own electric field called DON (dorsal octavolateral nucleus) [Bell et al. 2008]. For this essay I’m completely ignoring all these real complications and will simply use lateral line as a distance measurement.
The LL (lateral line) divides into two disjoint circuits, LL.a (anterior lateral line) covering the head before the ear, and LL.p (posterior lateral line) covering the trunk and tail behind the ear. LL.a connects to the hindbrain at R4 and LL.p connects at R6 [Chagnaud et al 2017]. The lateral line directly innervates the R.rs (reticulospinal) fast escape motor control neurons, including the paired giant Mauthner cells in R4 which trigger fast escape [Mirjany et al 2011].
Although hearing, vestibular, and lateral line use hair sensor sells, they are evolutionary independent [Chagnaud et al 2017]. Because both hearing and LL sense water oscillations, their signals overlap in frequency [Braun and Coombs 2000], with hearing sensing frequencies over 50Hz and LL sensing frequencies below 50Hz.
Lateral line circuits
Because this essay only uses LL for obstacle avoidance, it only implements the direct LL to R.rs connection, inhibiting movement toward an obstacle. The full LL circuit includes input to M.ic (inferior colliculus, hearing) and OT (optic tectum, blindsight) for higher-level navigation. The LL output to Hb.m (medial habenula) exists in the lamprey [Stephenson-Jones et al 2012] although possibly only the electrosensory LL and is used for thigmotaxis (wall hugging).
Lateral line circuit. Non-faded areas are implemented in the essay. CB.mon (medial octavolateral nucleus, cerebellum-like), Hb.m (medial habenula), LL (lateral line), M.ic (inferior colliculus / torus semicircular), MLR (midbrain locomotive region), N.sp (spinal motor neurons), R.rs (reticulospinal motor neurons), r4/r6 (hindbrain rhomobomeres 4 and 6).
To break down the circuit, first consider the fast escape Mauthner cell in r4 (hindbrain rhombomere 4), which connects directly to fast-twitch muscles. For an early vertebrate, a nearby predator could trigger the lateral line, which then triggered the Mauthner cell for a fast escape. The essay’s simulation uses the same circuit, but for obstacle avoidance instead of escape because I haven’t added any predators yet.
Next consider the thigmotaxis circuit using Hb.m (medial habenula) for gradient descent, where LL replaces the phototaxis light/dark gradient in essay 24 or chemotaxis odor gradient in essay 26. The lateral line gradient of water movement as the animal nears an obstacle drives Hb.m to balance wall avoidance with wall hugging. Note that the nAChR (nicotinic receptor) from R.ip (interpeduncular nucleus) to P.ldt (laterodorsal tegmental nucleus) is a goldilocks attract / avoid circuit [Wolfman et al 2018], which is the diagram’s Hb.m to MLR connection. At a distance, the weak glutamate-only R.ip to P.ldt signal is attractive, but nearby the stronger ACh amplification switches to aversion. The “nicotinic” is important here, because nicotine as a drug acts here for both nicotine’s attraction at low doses and its aversion at high doses. (Note: in this case “gradient descent” is the correct term, because the animal has a target distance so either too close or too far forms a signed error.)
Next consider the unmodulated LL to M.ic (inferior colliculus / torus semicircularis) to OT (optic tectum) circuit. Even ignoring optic input, OT has an egocentric map of sensory input, including the lateral line. Since the lateral line neuromasts are distributed across the animal’s body and head, each neuromast can give location-specific information about the distance to the obstacle or threat. The LL data becomes a marker on the OT map, enabling a more sophisticated OT navigation, where all the obstacles are marked on the map.
Finally consider the adaptive filter improvement from CB.mon (medial octavolateral nucleus) as a noise-cancelling system, where the noise is self-generated motion affecting the LL signal. CB.mon uses motor efference copies from MLR and B.rs and LL sensor input to predict the self-motion effect on LL. It sends the self-motion prediction to M.ic allowing M.ic to both filter out the self-motion, but also retain the unfiltered input. After filtering out self-motion, the remaining signal better represents the obstacles or threats. Although this signal could be called a prediction error, I think using filtered signal is a clearer and more useful description because the prediction itself is merely instrumental in producing a useful signal.
Cerebellum-like circuits
CB.mon (medial octavolateral nucleus) in the diagram above is a cerebellum-like system [Bell et al 2008], [Montgomery et al 2012]. Cerebellum-like systems are adaptive filters that remove predictable self-generated signals from self-motion. Because LL measures water motion but swimming also produces water motion, LL will primarily measure the animal’s own motion, unless the animal has paused. Since the self-motion would swamp the signal from obstacle, predators, or prey, CB.mon cleans up the signal by filtering out self-motion.
Social lateral line modulation
Modern fish are far more advanced animals than the simulation and famously swim in schools. The proximity of peer fish adds complexity to the lateral line. The fish needs to allow the peer to come nearer than a predator or even an obstacle.
Since lateral line is old, it’s unsurprising that schooling behavior is peptide-modulated. In zebrafish the neuropeptide pth2 reduces lateral line sensitivity and allows for closer schooling [Anneser et al 2020]. This effect is modulated by fish models of social anxiety, where anxious fish use larger distancing [Anneser et al 2022].