
The planned striatum model of essay 17 quickly runs into simulation problems because it’s missing priority selection between avoiding obstacles and seeking food. Obstacle avoidance needs a higher priority than seeking an odor plume, but a naive striatum doesn’t support that priority.
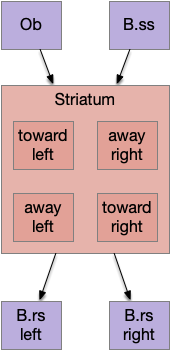
This model fails because this striatum has no priority of away (avoid) actions from toward (approach) actions. An animal can’t simply follow an odor blindly, ignoring obstacles, but this model doesn’t support that priority.
Tectum
Adding the tectum seems like the right solution, although I was planning on putting it off until dealing with vision.
The tectum (optic tectum / superior colliculus) is better known for its vision support, but the deeper tectum layers are a general action-decision system. At its lower levels near periaqueductal gray (M.pag) it has a topographic direction-based map on its intermediate level and an action-based map in the deep level.
The tectum and M.pag are neighbors, almost layers of each other, and in animals like the frog, the M.pag is as a deeper layer of the tectum.
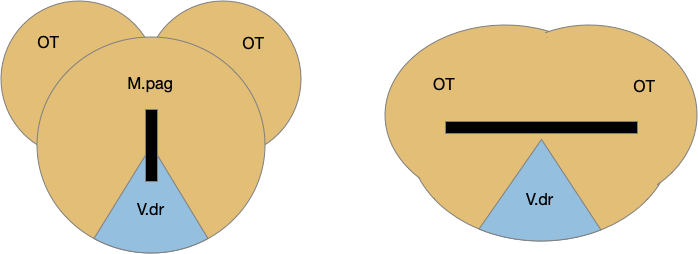
The tectum is an action organizer, not just a vision organizer. For the simulation, the action matters since the simulated animal doesn’t have vision.
Amphioxus, a non-vertebrate chordate that’s a model into pre-vertebrate evolution, has a few motor-related cells with the same genetic markers as the tectum [Pergner et al. 2020]. It’s conceivable that the amphioxus tectum is more action focused, since the amphioxus frontal eye is only a dozen photoreceptors with no lens.
Action categories
The tectum has split circuits for turning and for approach and avoid [Wheatcroft et al. 2022]. The simulation can use something like the following circuit.
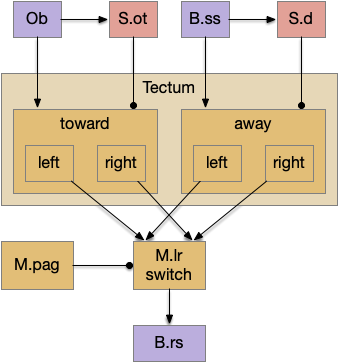
Approach (toward) senses like food odors excited toward actions, and avoidant (away) sense like touch excite away actions. Because the priority areas are split, each striatum can choose between non-priority options (left vs right). The priority resolves only later in the midbrain locomotor region, using context input to decide which major direction to use. In this split model, the simplified striatum circuit can work because all of striatum options are equal priority.
As a note on accuracy, the diagram misrepresents the actual olfactory path, specifically the real olfactory tubercle. In reality, olfaction has a distant, complicated path to the tectum.
Short-cut escape signal
The previous diagram is also misleading because it’s too organized, as if each function has a dedicated, planned circuit. Although the tectum itself is highly-organized, the downstream and modulating circuits are more ad hoc. For example, the zebrafish has an escape mechanism that short-cuts the tectum and drives the B.rs command motor directly [Zwaka et al. 2022].
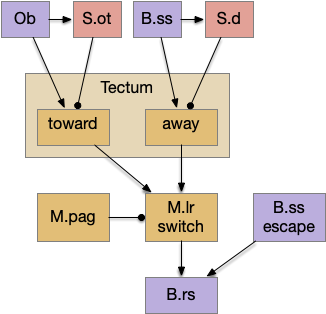
In the above diagram, the escape circuit short-circuits any decisions of the tectum and striatum. Relatedly, the “switch” area in M.lr isn’t as tidy as the diagram suggests. It’s more like that M.lr contains multiple actions which laterally inhibit each other in a priority scheme, modulated by M.pag.
As an additional correct, many of the modulators like M.pag affect the tectum directly, instead of the diagram’s dedicated priority-resolution function.
References
Pergner J, Vavrova A, Kozmikova I, Kozmik Z. Molecular Fingerprint of Amphioxus Frontal Eye Illuminates the Evolution of Homologous Cell Types in the Chordate Retina. Front Cell Dev Biol. 2020 Aug 4
Wheatcroft T, Saleem AB, Solomon SG. Functional Organisation of the Mouse Superior Colliculus. Front Neural Circuits. 2022 Apr 29
Zwaka H, McGinnis OJ, Pflitsch P, Prabha S, Mansinghka V, Engert F, Bolton AD. Visual object detection biases escape trajectories following acoustic startle in larval zebrafish. Curr Biol. 2022 Dec 5