
The previous proto-striatum model is flawed because it focused too much on sensory input and not enough on action efferent copies. To fix this focus, the model can use midbrain locomotive region (MLR) actions as a bias selector.
Recall that the simulation needed the striatum to solve an action jitter problem by introducing a win-stay bias. Once the animal turns left, it should bias toward continued left turns. Before the fix, the animal randomly chose a direction every 50ms, reversing itself, causing problems in avoiding corners and obstacles. The simulation problem was an action-selection problem not a sensor problem.
In the vertebrate striatum, action feedback comes from the MLR via the parafascicular thalamus (T.pf). The T.pf connection to the striatum is unique, both in its targeting of striatal interneurons (S.cin and S.pv), but also for its connection to the medium spiny projection neurons (S.spn), the main striatal neurons [Ragu et al. 2006]. T.pf connects directly to S.spn dendrites, not merely the spines as with other inputs. This direct connection potentially gives a stronger stimulus, and its uniqueness suggests it may be an older, more primitive connection.
Action-focused striatum model
So, I’m changing the striatum model to follow an action focus. After an action fires the motor command neurons (B.rs reticulospinal), the MRL sends an efferent copy of the motor command to the striatum via T.pf.
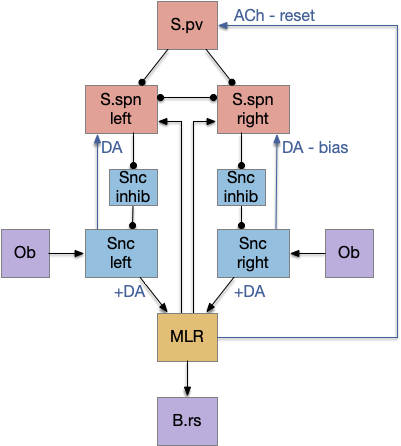
In the above diagram, the main sensor path is still from the olfactory bulb (Ob) to the substantia nigra pars compacta (Snc / posterior tuberculum) and then to MLR, basically a stimulus-response path. A previous action biases the sensory path for the next action by activating a corresponding S.spn, which disinhibits Snc, making the next sensory input more powerful.
Comparison with the previous model
As a comparison, the following diagram shows the previous striatal model. Unlike the new model, the final selected action didn’t bias the next action because there was no feedback connection. (The reset signal to S.pv is a different circuit, and doesn’t bias the decision because it applies to all choices equally.)
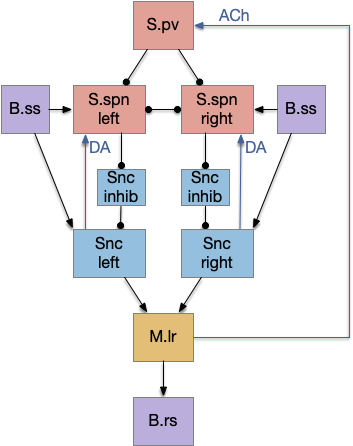
In addition, the sensory input must coordinate striatal disinhibition via S.spn with its excitation of the Snc action. Although not impossible evolutionarily, the double coordination required makes it less likely. The new model not only incorporates the action but simplifies the sensor circuit.
Parafascicular thalamus
For personal reference, here’s a summary of the T.pf connections [Smith et al. 2022].

Essentially all the T.pf inputs are motor efference copies and all the T.pf outputs are to the basal ganglia. Inputs include the following areas: vision/optic motor (OT and pretectum), midbrain locomotive region (MLR, M.pag, V.ppt, V.ldt), diencephalon locomotive region (H.zi), consummatory action (B.bp), forebrain attention (P.bf) and cortical action (C.fef, C.moss, C.gu). The cingulate cortex might be unusual (C.cc), although it also has motor areas.
Striatum as attention
Attention is a difficult topic, in part because it’s used in so many diverse ways that the word is often more confusing than helpful [Hommel et al. 2019], [Krauzlis et al. 2014]. However, I think it’s interesting that the action-based striatum model looks like selective attention.
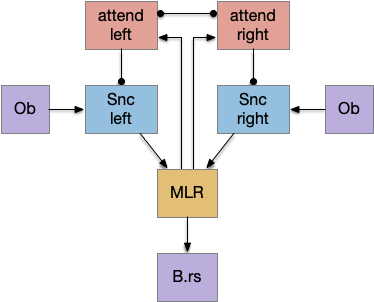
When a left action biases the next action to stay the same, its mechanism is to enhance the sensory path, as if it’s paying attention more to one side than another.
Engineering feedback: dopamine mistake
When implementing this idea, the simulation doesn’t need dopamine feedback. Instead of forcing the dopamine just because the basal ganglia has dopamine feedback I’m taking it out from the model. Since I’ve only implemented a prototype portion of the basal ganglia, this may be okay instead of a fatal flaw. When the full model arises, we’ll see if this is a mistake.
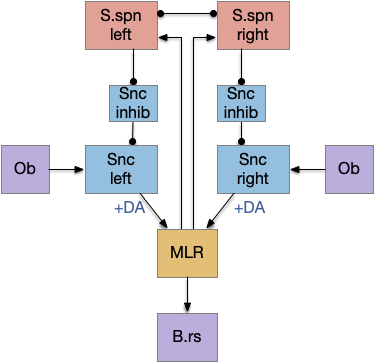
Notice that the only dopamine in this model is descending, with no ascending dopamine [Ryczko and Dubuc 2017].
References
Hommel B, Chapman CS, Cisek P, Neyedli HF, Song JH, Welsh TN. No one knows what attention is. Atten Percept Psychophys. 2019 Oct
Krauzlis RJ, Bollimunta A, Arcizet F, Wang L. Attention as an effect not a cause. Trends Cogn Sci. 2014 Sep;18(9):457-64
Raju DV, Shah DJ, Wright TM, Hall RA, Smith Y. Differential synaptology of vGluT2-containing thalamostriatal afferents between the patch and matrix compartments in rats. J Comp Neurol. 2006 Nov 10
Ryczko D, Dubuc R. Dopamine and the Brainstem Locomotor Networks: From Lamprey to Human. Front Neurosci. 2017 May 26
Smith JB, Smith Y, Venance L, Watson GDR. Thalamic Interactions With the Basal Ganglia: Thalamostriatal System and Beyond. Front Syst Neurosci. 2022 Mar 25