
Essay 13 is based on a study of zooplankton larva Platynereis as studied in [Conzelmann, 2011]. Specifically the apical sensory system and its control of the up and down swimming and sinking of a ciliary motor system. Importantly, the cilia motor system is very different from muscle systems.
Cilia cells operate independently, and can be organized by non-neural biomechanical systems, including gap junctions but also mechanical synchronization. In contrast, muscles need more accurate coordination for a sheet of muscle to contract together and for any opposing muscles to relax. A cilia-based system is simpler and potentially more primitive.
In addition to exploring cilia as a motor system, the Conzelmann study finds peptides from the apical tuft as control signals direct to the cilia ring, as opposed to neurotransmitters. (The larva does also have a traditional neurotransmitter system.)
Neuropeptides are much slower acting and longer lasting than neurotransmitters, and broadcast where neurotransmitters tend to direct to specific targets. Where a neurotransmitter might act in 10ms and disappear as quickly, a peptide might last for 10s of seconds or even minutes. As an illustration, knocking out the hypothalamic peptide orexin (by genetic manipulation), can cause narcolepsy in rats. One explanation for the narcolepsy is that orexin extends awakeness even with no stimulus for a minute or two, while the lack of orexin might allow sleep instantaneously when no stimulus is present.
Any sensory control of the cilia system is limited, restricted two two major variables. The cilia can wave faster or slower, or they can halt entirely. The cilia movement swims up in the water column, or sinks when the swimming stops. While there is a directional control toward light that uses photosensor near the cilia (using ACh), the one-dimensional apical control is the of the study and the essay.
The Conzelmann study found eleven distinct apical peptides, each tied to a specific sensor neuron. When they measured the effect on the cilia, nine of the eleven increased swimming rate. Five decreased arrest (stopping), while four had no effect on arrest rate. Two peptides both decreased swimming and increased the arrest rate.
The connection from the peptide sensors to the cilia appears to be direct, in contrast to muscle-based systems, which always have interneurons. The larva does have a separate central nervous system that develops into the adult nervous system, but the larva’s cilia nervous system appears to be entirely independent.
Evolutionary note: primitive or derived
Although the larva’s apical nervous system is simple, it may not be primitive, in the sense of the primitive nervous system of the oldest ancestor. It may have evolved separately as an optimization for the larval form, after the adult nervous system had evolved. Every living animal has hundreds of millions of years of evolution beyond the primitive animals, and co-evolution and devolution occur frequently. Because larva are necessarily small, vulnerable and released in large numbers, there may be a huge competitive pressure to optimize the simple forms, which may not reflect the primitive system.
The essay model of the zooplankton
The essay model follows the one-dimensional system. Sensors trigger neuropeptides, which trigger changes in the swim rate and arrest. It does not attempt to accurately model the fluid dynamics or even the sensors, but is only a simple system, where deeper locations have increased pressure and higher locations have increased light and heat.
The model is intended primarily as an executable thought experiment, not a solution. Creating a solution is more along the lines of control theory, which isn’t the aim of the exercise. Instead, it’s more designed to expose issues and choices of the model.
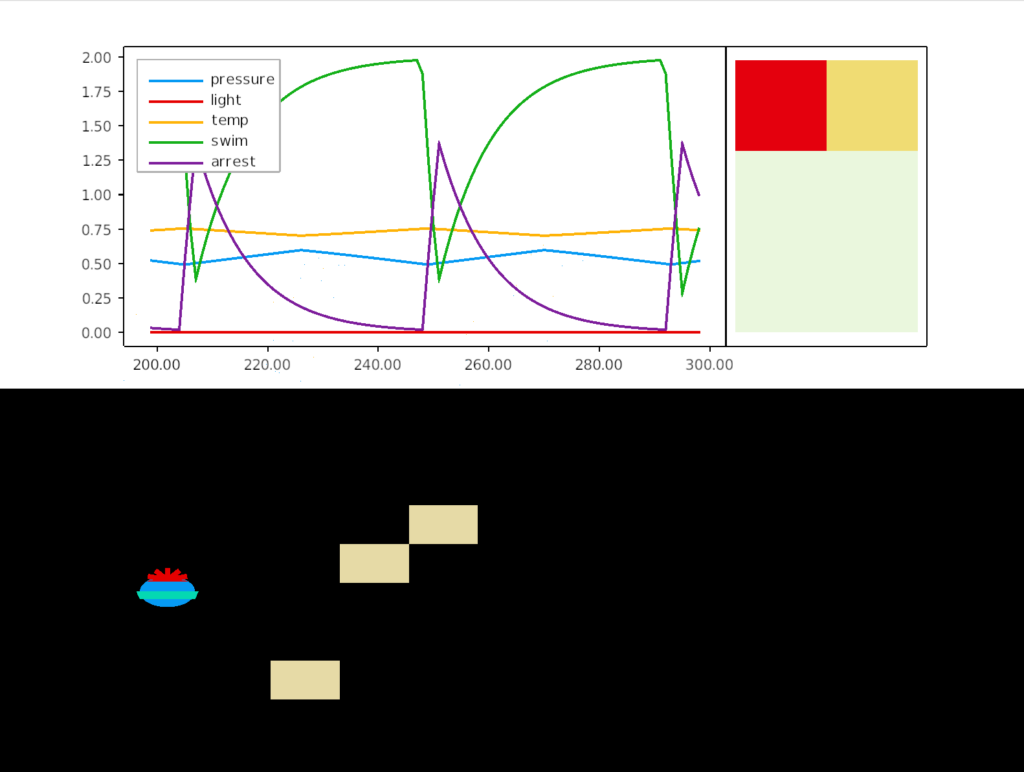
In this simulation, the larva oscillates between rising because of the pressure peptide, which increases swimming, until the rising triggers a too-hot sensor that releases a heat peptide, which triggers and arrest, which causes the larva to sink. The twin bounding limits of high pressure and high temperature keep the larva in a narrow band.
The meaning of a peptide
What does a peptide mean? Which seems like an odd question, but peptide communication has two ends, sender and receiver, which need to work together to express the behavior. There are several choices. A peptide could be sense-based: too-hot, high-pressure, light, hunger, predator, etc. Or it could represent a motor effect: increase swimming, or ciliary arrest. Possibly multiple peptides might express more subtle distinctions. Or the peptide might be an abstract modulation of the target cell: increase firing rate, or raise or lower trigger thresholds, without any specific sensory or action meaning to the peptide. [van den Poll, 2012] has an extensive discussion of how peptides appear in the nervous system.
Because the Conzelmann study found distinct peptides tied to senses, the essay model uses a sense-based system.
pub enum Peptide {
Pressure,
TempHot,
TempCold,
Light,
CO2
}An alternative model might focus on motor effects, where the sense neuron emits a peptide to drive a specific motor response:
pub enum Peptide {
SwimFaster,
SwimMedium,
Arrest
}This choice raises the question because evolution must create and coordinate both sides: the peptide selection, creation and release on the sending side, and peptide detection on the receiving side. The two mechanisms are genetically distinct. In fact, many neurotransmitters like dopamine (DA), serotonin (5HT), and acetylcholine (ACh), and multiple receptors with opposite or near-opposite effects. As anyone who’s been on a specification committed know, coordinating producers and consumers is a difficult and time-consuming task.
Raising the difficulty of understanding the system further, any meaning of a particular peptide doesn’t necessarily follow a human-friendly concept because evolution isn’t a human designer. At very least, a peptide is likely to have multiple idiosyncratic uses that more resemble cheap hacks than careful design.
All-or-nothing vs graded peptides
The essay model chooses an all-or-nothing behavior as opposed to a graded response. The sensory neurons don’t release any peptides until the sense crosses a threshold.
fn sense_hot(body: Res<Body>, mut peptides: OutEvent<Peptide>) {
if body.temperature() > 0.75 {
peptides.send(Peptide::TempHot)
}
}An alternative model might choose a graded response:
if temp > 0.65 {
peptides.send(Peptide::TempHot(temp - 0.65));
}Which is the better model isn’t clear. The graded response appears more precise, but the the precision isn’t needed for the behavior, it might just be wasted effort, because producing and releasing peptides has a cost. If low levels aren’t going to have an effect for temp=0.66, then producing a tiny amount of peptides that will be ignored is wasted effort.
In this scenario, the larva achieves its primary goals — avoiding excess heat by going too high and avoiding dropping too low — without needing extra precision. Where a control-theory designed system would be more optimal, the extra effort might not be worth the extra complication.
Minutes not milliseconds
In this scenario, minutes is an exaggeration, but seconds vs milliseconds is still a side time difference. Is there an advantage or disadvantage to use peptides with a long time scale, or is the time difference irrelevant to the scenario? Even without the timing difference, peptides might still have an advantage of multiple codes for specific senses, allowing complex behavior without needing complicated wiring. For a zooplankton larva, there might be a premium for simplicity.
The model for the delay looks like the following:
peptides = peptides * 0.9 + peptide_input;where peptides is a vector and the decay constant reflects the ticking simulation, not the decay of any particular peptide. In theory, the decay should differ with each peptide.
The decay rate does affect the performance of the model, particularly the oscillation. There’s an entire field of control theory that studies these effects and how to optimize them. So, the different matters in the detail. But the main behavior of avoiding heat while staying near the surface doesn’t differ much.
Consider chemotaxis (moving toward or away from a scent.) An odor sensor might only receive a scent molecule every second, and randomly distributed, which is a problem if the animal forgets everything every 20ms. With a fast neurotransmitter, the animal would forget its potential prey or ignore a predator after a second or two of fleeing. In that situation, using a peptide to keep track of prey or predator, keeping consistent motivation for a minute, might have a huge different. But a simulation model that is too consistent, simple, or lacking statistical variation might miss the need for a longer timeframe peptide. But this specific scenario only measures slow, consistent variables like pressure, heat and light. So the extended timescale of peptides may not make a behaviorally significant difference.
Discussion
I think the value of this essay is just doing an exercise to try the peptide model as opposed to the standard connective model. The real value might appear in a later essay, where a relatively simple peptide model that handles much of motivational state interacts with a more conventional connective model for motor control and behavior.
References
Conzelmann M, Offenburger SL, Asadulina A, Keller T, Münch TA, Jékely G. Neuropeptides regulate swimming depth of Platynereis larvae. Proc Natl Acad Sci U S A. 2011 Nov 15;108(46):E1174-83. doi:10.1073/pnas.1109085108. Epub 2011 Oct 17.
van den Pol AN. Neuropeptide transmission in brain circuits. Neuron. 2012 Oct 4;76(1):98-115. doi: 10.1016/j.neuron.2012.09.014. PMID: 23040809; PMCID: PMC3918222.