
The simple phototaxis implementation exposes a few problem with the simulation, both from running it and from reviewing neuroscience to critique it.
Interrupts
The essay doesn’t currently implement any interrupt mechanism. When running into darkness, the animal turns around and starts an area restricted search (ARS), but if the animal is in the middle of a long Levy path, it will cross the border and the search will not find the border.
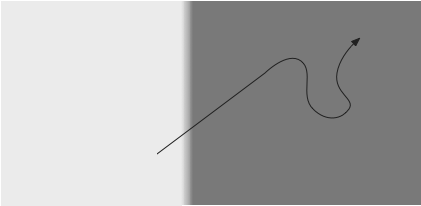
The problem here is that the ARS starts too late because the animal doesn’t interrupt the current behavior when encountering the border.
One solution is to create an interrupt (orientation) system, which exists in the vertebrate brain in V.ppt (peduncular pontine nucleus), and uses ACh (acetylcholine) to interrupt the current behavior. A natural location for the interrupt is V.ppt for the signal and the stratum as the plan representation, interruptible via ACh interrupts to the striatum.
Another solution is to avoid the uninterruptible behavior entirely, where the problem is the essay’s Levy walk implementation. The essay pre-computes the length of a run instead of continuously creating extensions. In contrast the zebrafish larva swims in bouts, but longer runs are made of multiple forward bouts.
Zebrafish random walk (ARTR area)
The essay’s random walk does not match actual zebrafish search behavior. The essay uses a turn-and-run model where the turn and run length are computed randomly. The zebrafish has a hindbrain oscillator, the ARTR (anterior rhombencephalic turning region) which selects left and right turns [Karpenko et al 2020].
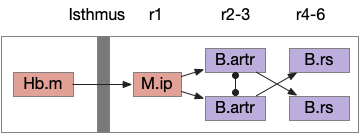
In zebrafish, turns and runs are selected independently and can be chained differently. Instead of turn-run-turn-run as in the essay, the zebrafish can have turn-turn or run-run patters. Zebrafish turn direction is also correlated, as opposed to the random walk’s turn independence. A zebrafish left turn is more likely to follow a left turn.
When encountering darkness, the same-direction turns increase. When encountering light, alternating turns increase. Together with the sharp turn (O-bend) followed b shallower turns, this behavior should create a spiral-like search for the light area.
Note: this specialized circuitry in the hindbrain suggests that random search is a primitive behavior. Although the essay put the Levy walk logic in the midbrain, it belongs in the more primitive hindbrain.
Head direction
[Petrucco et al 2023] report that M.ip is highly connected with head direction axons from B.dtg (dorsal tegmental area of Gudden). This head direction does not receive vestibular input, but is likely derived from motor efferent copies. Since both M.ip and B.dtg are r1-derived regions and possibly ante-vestibular, this head-direction and M.ip connection may be ancient.
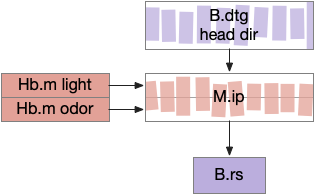
This organization is strikingly similar to the fruit fly’s ellipsoid body (EB), protocerebral bridge (PB), and fan-shaped body (FB) in the central complex (CX) [Hulse et al 2021]. EB and PB calculate head direction. The fan-shaped body merges head direction with goal direction to produce motor commands. In this diagram, M.ip represented as if it resembles the fan-shaped body.
If the M.ip functionality is similar to the fan-shaped body, it’s highly likely to be convergent evolution, not homology because amphioxus lacks any similar structure.
Dark search
When zebrafish are plunged into darkness, they initiate a search that continues for about five minutes. The darkness behavior increases speed and straight behavior [Horstick et al 2017]. In other words, phototaxis is not just gradient behavior but also has steady-state darkness behavior. Because zebrafish require light to hunt, darkness in itself is is an area to avoid.
The essay is purely gradient based and has no speed changes. Photokinesis is moving faster in darkness and slower in light, which will bias the time spent in the light area.
References
Hulse, B. K., Haberkern, H., Franconville, R., Turner-Evans, D., Takemura, S. Y., Wolff, T., … & Jayaraman, V. (2021). A connectome of the Drosophila central complex reveals network motifs suitable for flexible navigation and context-dependent action selection. Elife, 10.
Horstick EJ, Mueller T, Burgess HA. Motivated state control in larval zebrafish: behavioral paradigms and anatomical substrates. J Neurogenet. 2016 Jun;30(2):122-32.
Karpenko S, Wolf S, Lafaye J, Le Goc G, Panier T, Bormuth V, Candelier R, Debrégeas G. From behavior to circuit modeling of light-seeking navigation in zebrafish larvae. Elife. 2020 Jan 2;9:e52882.
Petrucco L, Lavian H, Wu YK, Svara F, Štih V, Portugues R. Neural dynamics and architecture of the heading direction circuit in zebrafish. Nat Neurosci. 2023 May;26(5):765-773. doi: 10.1038/s41593-023-01308-5.