[Arendt 2015] examines the fundamental divisions of the nervous system by looking at ancestral cell divisions in some of the earliest animals, specifically a multicellular amoeboid bottom-feeder that glides on a mucus foot like a slug. The archaoslug moves like an amoeba instead of a true slug because it’s body isn’t symmetrical bilateral: there’s no front or side. The Adrent study is interesting for the essays because it fundamentally splits chemosensory control (hypothalamic and olfactory) split from mechanosensory / optosensory sense and muscle brain at the cell type and developmental level.
In this proto-slug, cells have specialized into three major classes:
External skin with mechanosensory and optosensory cells
Internal digestive gut
Mucociliary sole with chemosensory and locomotion cells
The mucociliary sole moves with cilia gliding over mucus. Chemical sensors that detect food choose when to stop. A similar locomotive strategy is described in [Smith 2015] and [Senatore 2017] for the existing algae-grazing, disc-shaped animal Trichoplax, which lacks any nerves at all and only has six cells in total [Smith 2014].
Locomotion and food searching for the archaoslug is simple: stop when the chemosensors detects food (algae), and move in a random brownian direction when no food is available. A simple chemical sensor and a peptide-based broadcast system would suffice, as in Trichoplax. Because the bacterial mats may have dominated the Precambrian environment, the brownian motion pausing for food could work.
The ventral skin specialized into mechanosensory cells, optosensory cells, and contractile cells which developed into the first neurons and muscles. The animal can avoid obstacles and threats using nerve nets that broadcast and repeat signals, like the repeating nerve nets in cnidaria (jellyfish, sea anemone, and corals) [Seipel 2005]. Note, though, the control circuits for between locomotion (mucociliary sole) and muscles (skin/body contractions) are distinct, and don’t even coordinate. A sea anemone or a slug will contract when touched, but the sea anemone has no locomotion and the slug’s contraction isn’t its primary locomotion. Similarly, an archeoslug with primitive muscles might only use the muscles to avoid obstacles, contracting when it runs into something, but its primary motion remains the ciliary, non-muscular sole. Meaning, the locomotive drive (arrest, approach, avoid) is controlled independently from navigation (spatial obstacle avoidance.)
This division into three types influences all later cell development, because the initial decisions shape the later evolved cell types. Genetic signaling to choose between the three might have created a path dependent split between three types.
Discussion
This split between chemosensory sole and mechano- and opto-sensory skin and muscle obviously mirrors the vertebrate split between the limbic system (olfactory and hypothalamic) and optic tectum system, but with a different spin. The limbic system is generally described as a motivational and emotional center. The root word for both motivation and emotion is the Latin movere, to move, which has less baggage than either word. The mucociliary sole area does move and control movement, but it’s hardly an emotional center. But the mucociliary sole area isn’t unique in its control of motion, because the unrelated skin/muscle area controls navigation.
Treating the mind as independent, conflicting centers resembles Dawkins’ descriptions of genes in The Selfish Gene, where each gene works independently and in competition with others, and coordination only occurs for mutual benefit. The general idea of competing mental centers is also in Minsky’s Society of mind, and the idea is older than either. So, the value of the archaeoslug isn’t the general idea of a divided mind, but the specific division that occurred in evolution.
References
Arendt D, Benito-Gutierrez E, Brunet T, Marlow H. Gastric pouches and the mucociliary sole: setting the stage for nervous system evolution. Philos Trans R Soc Lond B Biol Sci. 2015 Dec 19;370(1684):20150286. doi: 10.1098/rstb.2015.0286. PMID: 26554050; PMCID: PMC4650134.
Dawkins, Richard. The Selfish Gene. Oxford University Press, 2006.
Minsky, Marvin. Society of mind. Simon and Schuster, 1988.
Senatore A, Reese TS, Smith CL. Neuropeptidergic integration of behavior in Trichoplax adhaerens, an animal without synapses. J Exp Biol. 2017 Sep 15;220(Pt 18):3381-3390. doi: 10.1242/jeb.162396. PMID: 28931721; PMCID: PMC5612019
Smith CL, Pivovarova N, Reese TS. Coordinated Feeding Behavior in Trichoplax, an Animal without Synapses. PLoS One. 2015 Sep 2;10(9):e0136098. doi: 10.1371/journal.pone.0136098. PMID: 26333190; PMCID: PMC4558020.
Smith CL, Varoqueaux F, Kittelmann M, Azzam RN, Cooper B, Winters CA, Eitel M, Fasshauer D, Reese TS. Novel cell types, neurosecretory cells, and body plan of the early-diverging metazoan Trichoplax adhaerens. Curr Biol. 2014 Jul 21;24(14):1565-1572. doi: 10.1016/j.cub.2014.05.046. Epub 2014 Jun 19. PMID: 24954051; PMCID: PMC4128346.
Essay 13 is based on a study of zooplankton larva Platynereis as studied in [Conzelmann, 2011]. Specifically the apical sensory system and its control of the up and down swimming and sinking of a ciliary motor system. Importantly, the cilia motor system is very different from muscle systems.
Cilia cells operate independently, and can be organized by non-neural biomechanical systems, including gap junctions but also mechanical synchronization. In contrast, muscles need more accurate coordination for a sheet of muscle to contract together and for any opposing muscles to relax. A cilia-based system is simpler and potentially more primitive.
In addition to exploring cilia as a motor system, the Conzelmann study finds peptides from the apical tuft as control signals direct to the cilia ring, as opposed to neurotransmitters. (The larva does also have a traditional neurotransmitter system.)
Neuropeptides are much slower acting and longer lasting than neurotransmitters, and broadcast where neurotransmitters tend to direct to specific targets. Where a neurotransmitter might act in 10ms and disappear as quickly, a peptide might last for 10s of seconds or even minutes. As an illustration, knocking out the hypothalamic peptide orexin (by genetic manipulation), can cause narcolepsy in rats. One explanation for the narcolepsy is that orexin extends awakeness even with no stimulus for a minute or two, while the lack of orexin might allow sleep instantaneously when no stimulus is present.
Any sensory control of the cilia system is limited, restricted two two major variables. The cilia can wave faster or slower, or they can halt entirely. The cilia movement swims up in the water column, or sinks when the swimming stops. While there is a directional control toward light that uses photosensor near the cilia (using ACh), the one-dimensional apical control is the of the study and the essay.
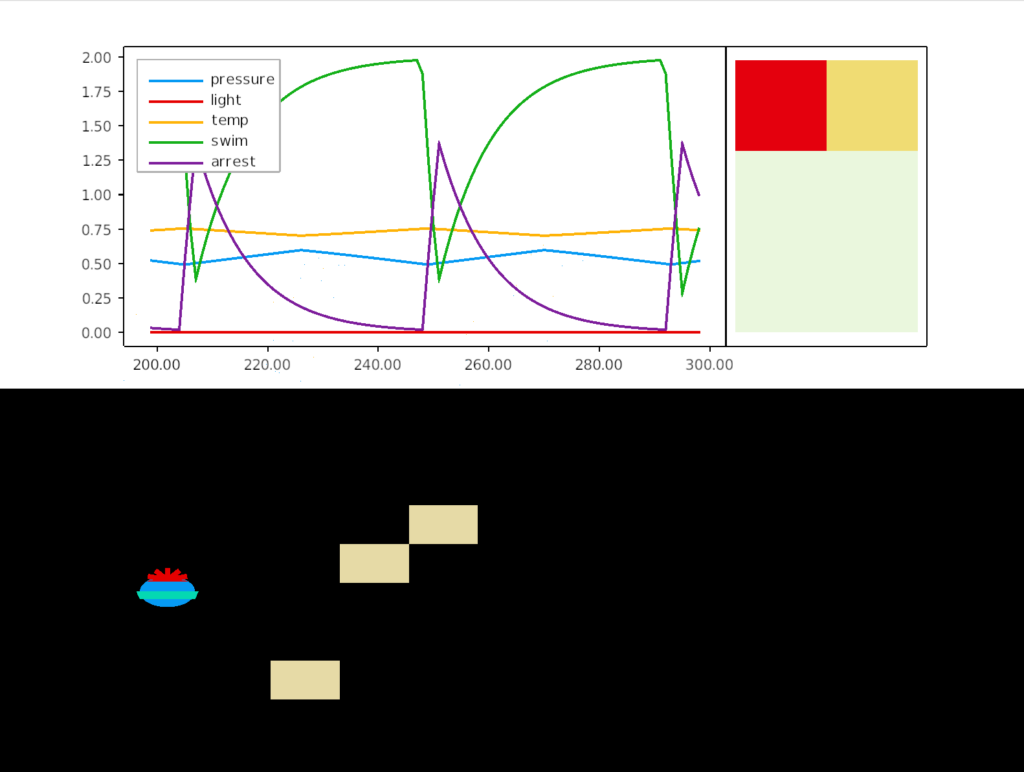
The Conzelmann study found eleven distinct apical peptides, each tied to a specific sensor neuron. When they measured the effect on the cilia, nine of the eleven increased swimming rate. Five decreased arrest (stopping), while four had no effect on arrest rate. Two peptides both decreased swimming and increased the arrest rate.
The connection from the peptide sensors to the cilia appears to be direct, in contrast to muscle-based systems, which always have interneurons. The larva does have a separate central nervous system that develops into the adult nervous system, but the larva’s cilia nervous system appears to be entirely independent.
Evolutionary note: primitive or derived
Although the larva’s apical nervous system is simple, it may not be primitive, in the sense of the primitive nervous system of the oldest ancestor. It may have evolved separately as an optimization for the larval form, after the adult nervous system had evolved. Every living animal has hundreds of millions of years of evolution beyond the primitive animals, and co-evolution and devolution occur frequently. Because larva are necessarily small, vulnerable and released in large numbers, there may be a huge competitive pressure to optimize the simple forms, which may not reflect the primitive system.
The essay model of the zooplankton
The essay model follows the one-dimensional system. Sensors trigger neuropeptides, which trigger changes in the swim rate and arrest. It does not attempt to accurately model the fluid dynamics or even the sensors, but is only a simple system, where deeper locations have increased pressure and higher locations have increased light and heat.
The model is intended primarily as an executable thought experiment, not a solution. Creating a solution is more along the lines of control theory, which isn’t the aim of the exercise. Instead, it’s more designed to expose issues and choices of the model.
In this simulation, the larva oscillates between rising because of the pressure peptide, which increases swimming, until the rising triggers a too-hot sensor that releases a heat peptide, which triggers and arrest, which causes the larva to sink. The twin bounding limits of high pressure and high temperature keep the larva in a narrow band.
The meaning of a peptide
What does a peptide mean? Which seems like an odd question, but peptide communication has two ends, sender and receiver, which need to work together to express the behavior. There are several choices. A peptide could be sense-based: too-hot, high-pressure, light, hunger, predator, etc. Or it could represent a motor effect: increase swimming, or ciliary arrest. Possibly multiple peptides might express more subtle distinctions. Or the peptide might be an abstract modulation of the target cell: increase firing rate, or raise or lower trigger thresholds, without any specific sensory or action meaning to the peptide. [van den Poll, 2012] has an extensive discussion of how peptides appear in the nervous system.
Because the Conzelmann study found distinct peptides tied to senses, the essay model uses a sense-based system.
pub enum Peptide {
Pressure,
TempHot,
TempCold,
Light,
CO2
}
An alternative model might focus on motor effects, where the sense neuron emits a peptide to drive a specific motor response:
This choice raises the question because evolution must create and coordinate both sides: the peptide selection, creation and release on the sending side, and peptide detection on the receiving side. The two mechanisms are genetically distinct. In fact, many neurotransmitters like dopamine (DA), serotonin (5HT), and acetylcholine (ACh), and multiple receptors with opposite or near-opposite effects. As anyone who’s been on a specification committed know, coordinating producers and consumers is a difficult and time-consuming task.
Raising the difficulty of understanding the system further, any meaning of a particular peptide doesn’t necessarily follow a human-friendly concept because evolution isn’t a human designer. At very least, a peptide is likely to have multiple idiosyncratic uses that more resemble cheap hacks than careful design.
All-or-nothing vs graded peptides
The essay model chooses an all-or-nothing behavior as opposed to a graded response. The sensory neurons don’t release any peptides until the sense crosses a threshold.
An alternative model might choose a graded response:
if temp > 0.65 {
peptides.send(Peptide::TempHot(temp - 0.65));
}
Which is the better model isn’t clear. The graded response appears more precise, but the the precision isn’t needed for the behavior, it might just be wasted effort, because producing and releasing peptides has a cost. If low levels aren’t going to have an effect for temp=0.66, then producing a tiny amount of peptides that will be ignored is wasted effort.
In this scenario, the larva achieves its primary goals — avoiding excess heat by going too high and avoiding dropping too low — without needing extra precision. Where a control-theory designed system would be more optimal, the extra effort might not be worth the extra complication.
Minutes not milliseconds
In this scenario, minutes is an exaggeration, but seconds vs milliseconds is still a side time difference. Is there an advantage or disadvantage to use peptides with a long time scale, or is the time difference irrelevant to the scenario? Even without the timing difference, peptides might still have an advantage of multiple codes for specific senses, allowing complex behavior without needing complicated wiring. For a zooplankton larva, there might be a premium for simplicity.
The model for the delay looks like the following:
peptides = peptides * 0.9 + peptide_input;
where peptides is a vector and the decay constant reflects the ticking simulation, not the decay of any particular peptide. In theory, the decay should differ with each peptide.
The decay rate does affect the performance of the model, particularly the oscillation. There’s an entire field of control theory that studies these effects and how to optimize them. So, the different matters in the detail. But the main behavior of avoiding heat while staying near the surface doesn’t differ much.
Consider chemotaxis (moving toward or away from a scent.) An odor sensor might only receive a scent molecule every second, and randomly distributed, which is a problem if the animal forgets everything every 20ms. With a fast neurotransmitter, the animal would forget its potential prey or ignore a predator after a second or two of fleeing. In that situation, using a peptide to keep track of prey or predator, keeping consistent motivation for a minute, might have a huge different. But a simulation model that is too consistent, simple, or lacking statistical variation might miss the need for a longer timeframe peptide. But this specific scenario only measures slow, consistent variables like pressure, heat and light. So the extended timescale of peptides may not make a behaviorally significant difference.
Discussion
I think the value of this essay is just doing an exercise to try the peptide model as opposed to the standard connective model. The real value might appear in a later essay, where a relatively simple peptide model that handles much of motivational state interacts with a more conventional connective model for motor control and behavior.
References
Conzelmann M, Offenburger SL, Asadulina A, Keller T, Münch TA, Jékely G. Neuropeptides regulate swimming depth of Platynereis larvae. Proc Natl Acad Sci U S A. 2011 Nov 15;108(46):E1174-83. doi:10.1073/pnas.1109085108. Epub 2011 Oct 17.
van den Pol AN. Neuropeptide transmission in brain circuits. Neuron. 2012 Oct 4;76(1):98-115. doi: 10.1016/j.neuron.2012.09.014. PMID: 23040809; PMCID: PMC3918222.
My initial idea for essay 13 is to explore the chimaeral model [Tosches, 2013], which models the bilaterial mind as composed of two poles, an apical pole around chemosensory, peptides and possibly a more broadcast model, and a motor pole around central pattern generators and muscle control.
The apical pole is based on the larvae of many marine animals, zooplankton. Platynereis for example is a marine worm with a ciliated larva that follows the apical plan. The cilia is important, because cilia and muscles have very different control needs. Cilia can work largely independently, or in a self-organizing fashion where neighboring cells beat together. Any control system is intrinsically modulating, not directly controlling. In contrast, muscles require a nerve net to function, because of the need to coordinate the contraction of many muscle cells.
I’m hoping that exploring the apical model of modulating downstream systems will also counterbalance the natural inclination to imagine neural control as a micromanager controlling every individual muscle or gland, or if not micromanaging, then delegating in a strict naive hierarchical fashion.
A question with such a simple animal with behavior that’s easily programmed, is what’s the value of a programming model, when it’s easily solved? Or, perhaps better, what kinds of models might be useful?
This problem is particularly an issue for an essay model, because it has more resemblance to old-style AI, which runs into the problem of wishful mnemonics [McDermott 1976], where the naming of variables and functions substitutes for any actual insight.
For example, some future essay might model hunger in the hypothalamus:
But all the work here is in the naming: goals, motivation, even the base “hunger,” which is simply a floating point number. The model itself is trivial; all the work is in the illusion given by pointed naming. McDermott suggests replacing all variables with generated symbols like G0019, and seeing if the model still works.
So, where is the value? Not as a solution, because the solution is trivial. But I think the model can have value as an executable thought experiment, like a diagram, not as a solution. The advantage of a program is that it can catch handwaving assumptions because you have to make decisions to write a program.
Programming infrastructure: ECS and ticks
The essay models use a loosely coupled system with a tick by tick simulation. The loose coupling forces communication to be an important issue, because neural fibers and connectivity is as important as grey matter. The ticking simulation forces time to be a fundamental issue.
For a base engine, the essays use an ECS (entity, component, system) based on the Bevy game engine. Each tick runs all the systems, which each run independently, using multiple threads where possible.
A setup phase registers the systems with the ECS engine. For example, a pressure-sensing neuron in the zooplankton might register with the application as follows, where sense_pressure is a function:
app.system(Update, sense_pressure);
The simulation proceeds with ticks to execute all the systems, called as follows:
app.tick();
Because of the loose coupling, the model isn’t a single algorithm but a collection of independent systems that each influence the whole.
Here the body represents the physical model, so the sensory neuron is receiving pressure from the POV of the body, as opposed to knowing about the broader world. If it senses sufficient water pressure, it broadcasts a peptide as a message.
Note that the sensor neuron doesn’t know what the peptide will do. Although in the full model, the pressure peptide will cause the cilia to beat faster, swimming up, that knowledge is outside of the sensor.
In this example, the pressure sensor is a unique neuron so there’s no need to have an individual component representing it, but a more complicated animal would have specific components for each physical sensor.
Tosches, Maria Antonietta, and Detlev Arendt. “The bilaterian forebrain: an evolutionary chimaera.” Current opinion in neurobiology 23.6 (2013): 1080-1089.
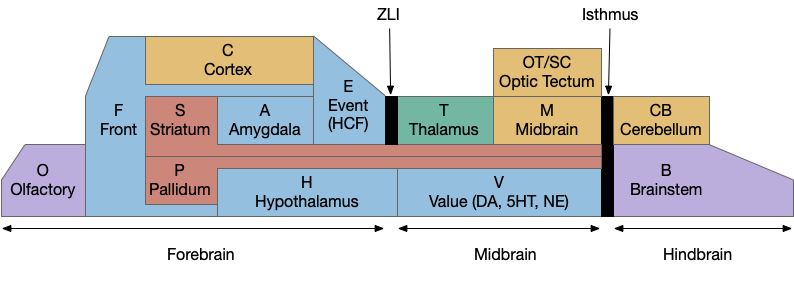
Because learning and remembering the vast number of vertebrate brain nuclei and areas and connections is a daunting task, I’ve needed to use a simplified mnemonic model to make some sense of the whole and show how individual pieces with together. The following is a simplified model of the brain, emphasizing subcortical areas, and with distinct initial letters so a quick scan of an abbreviation is pre-categorized in its general location.
Disclaimer: this is a personal model and may not accurately reflect any current consensus of neuroscience. Specifically, the abbreviations are different from the common neuroscience appreciations.
A key decision of the model is to use consistent single-letter prefixes. Some of these letters are clear, like “H” for hypothalamus, “A” for amygdala, and “T” for thalamus, but others are a bigger stretch. The “V” for monoamines (dopamine, serotonin, norepineprine) and acetylcholine (ACh) is meant to indicate “value” reflecting their roles in providing operating current values. The hippocampal formation, which is a center of episodic memory, uses “E” for event or episodic because “H” for hypothalamus is already taken.
Not every area fits cleanly in a box, partly because evolution fails to follow clear, human-friendly boundaries. But also because several areas are intrinsically liminal: their function depends on manipulating data from multiple areas. For example, the cingulate cortex (C.cc or F.ac), which has functions related to attention and conflict detection, is often placed with the frontal area as part of the prefrontal cortex (PFC, or F.pfc), but also ranges along the cortex to the hippocampus. The zona incerta (H.zi), which is also attention-related, is strongly related to the midbrain motor areas and the optic tectum, despite being physically adjacent to, or possibly part of the hypothalamus. The pre optic areas (P.poa or H.poa), which are related to exploration, reproductive behavior, and aggression, have traditionally been grouped with the hypothalamus (H), but more recently grouped with pallidal (P) areas.
The bilaterian chimeral brain (“H” and “OT”)
The bilaterian chimeral forebrain theory [Tosches and Arendt, 2013] is an interesting organizational principle that the mnemonic model uses to place the hypothalamus (H) firmly in the forebrain as a central organizing pole and the midbrain motor areas centered around the optic tectum/superior colliculus (OT). The coloring scheme of blue and brown is meant to group areas with their primary pole, motivational and limit areas with the hypothalamus and motor and habit cortex with the midbrain and optic tectum.
The chimeral model itself suggests that the bilaterian brain, including insects and vertebrates, is derived from two ancestral sources, the forebrain from the larval apical tuft and the midbrain/hindbrain from the adult motor areas. Larvae from mollusks and annelids still show the ancestral apical areas. The apical tuft is a chemosensory area that communicates using broadcast neuropeptides, while the bilaterian motor areas use connected neurons and neurotransmitters. The hypothalamus with its melange of over 100 neuropeptides, internal chemical sensing, and connections with visceral and olfactory areas is a descendent of the apical tuft. Contrariwise, the central pattern generators of the spinal cord, hindbrain and midbrain would be descendants of the adult motor areas.
The diencephalon and prosomeric model (“H” and “T”)
The mnemonic model groups everything between the zona limitans intrathalamica (ZLI) and the isthmus organizer into the midbrain and everything rostral to it into the forebrain, including the hypothalamus, the zona incerta (H.zi) and the pre-thalamic eminence (P.em), while leaving the thalamic reticular nucleus (T.r) with the thalamus, but that entire region is currently under debate by neuroscientists (and the model doesn’t reflect any of the theories directly.)
The diencephalon is an older anatomical term that groups the pre-tectal areas (M.pt), the thalamus (T), the hypothalamus (H) and the zone between H and T (the “pre-thalamus”) into a single region. The diversity of function and of developmental gene expression have raised the question of the usefulness of that grouping, in particular putting the hypothalamus and thalamus into a shared group.
The prosomeric model [Puelles and Rubinstein, 1993] instead splits the hypothalamus and thalamus into two regions made of five sub-regions, p1-p3 representing the pre-tectal, thalamic, pre-thalamic areas, and hy1 and hy2 representing two regions of the hypothalamus. p3 includes interesting liminal areas zona incerta (H.zi), pre-thalamic eminence (P.em), and T.r.
But critics of the prosomeric model like [Bedont, 2015] have argued that some areas of the pre-thalamus (prosomeric p3) are closely genetically and developmentally tied with the hypothalamus, not the thalamus, questioning whether prosomere is a useful organizing principle.
That area of the brain has an unusual genetic and chemical marker at the ZLI, where the Shh gene expression rises from its usual basal location to the top of the embryo, creating a thin chemical boundary between the thalamus and pre-thalamus [Kiecker and Lumsden, 2004]. The ZLI splits both the diencephalon model and the prosomeric model in two.
For simplicity, the mnemonic model uses ZLI as a dividing line and since the model is not detailed enough to give p3 its own box, it can remain neutral on the neuroscience debate, although it does follow the prosomeric model enough to strongly split the hypothalamus from the thalamus and also put H with the forebrain.
Striatum and pallidum (“S” and “P”)
All areas with striatal or pallidal structure are grouped together, essentially everything in the sub-pallium derived from the median ganglionic eminence (MGE) and lateral ganglionic eminence (LGE), following the model of [Swanson 2000.]
The stratum and pallidum (S/P, basal ganglia or sub-pallium) are interested because they’re key components of conditioned stimuli, conditioned responses, conditioned values, reinforcement learning, feedbacks loop in the cortex, and communication between the cortex and the midbrain motor areas (M and OT) and motivational areas (H and V) as well as combining episodic context (E.hc, hippocampus) and motivational context (A, amygdala). As well as communicating between areas, the striatum (S/P) is a key component of communication within the cortex and frontal areas with cortical intratelencephalic neurons (IT, C.5a) strongly connecting to the striatum.
The grand simplification lumps many areas into one naming group. In the traditional striatum, the caudate and putamen becomes S.cp (or. S.d for dorsal striatum). The nucleus accumbens becomes S.v or S.nac, and if greater detail is needed, medial shell (S.msh) and core (S.core).
The traditional pallidum becomes P.ge and P.gi for internal and external globus pallidus, but can be more specific for the habenular-projecting neurons (P.h), and the endopeduncular nucleus, which is a habenular-projecting area for mice becomes (P.epn). The ventral pallidum is P.v (or P.si if the distinction is needed.)
Since the septum also derives primarily from subpallial sources, it becomes S.e and P.e (S/P related to “E” hippocampus) or S.ls and P.ms for lateral and medial septum (as well as P.ts, P.sf, and P.bac for posterior septum and P.db or P.msdb for the diagonal band.)
Since the extended amygdala includes striatal and pallidal components, becomes S.a and P.a for the amygdala S/P (or S.cea, P.bst for central amygdala and bed nucleus of the stria terminals.)
If the pre-optic areas (H.poa) are instead grouped with the pallidum, they become P.poa (or more specifically P.mpo, P.lpo.)
And, since the pre-thalamic eminence is the major source for pallidal cells projecting to the habenula (P.h to Hb), it receives an honorary P.em despite not technically belonging to the telencephalon.
In theory, following the mnemonic model would put the output of the basal ganglia as M.snr and V.snc because the substantia nigra pars reticulara is both in the midbrain motor area (M) and projects to it and the substantia nigra pars compacta is part of the dopamine circuit in the V area, but the initial S gives a natural give S.nr and S.nc.
If there is anyone who has read all through that list, I hope there’s some understanding of why it’s useful to initially lump all the above into S and P groups in the process of learning all the names.
Hippocampal formation (“E”)
Although the initial letter “E” is the least effective, grouping hippocampal formation areas is helpful despite the problems. The hippocampus itself becomes E.hc, dentate gyrus E.dg, ca1 is E.ca1, the subiculum is E.sub, the lateral and medial entorhinal cortices become E.lec and E.mec, and the prerhinal and parahippocampal are E.pr and E.phc (or E.por for the equivalent postrhinal cortex in rodents.)
Thalamus (collo- and lemno-)
One interesting division in the thalamus that the model does not portray is the [Butler 2008] distinction between collothalamic and lemnothalamic areas, where “collo” refers to the superior and inferior colliculi, essentially the optic tectal (OT) area, and “lemno” are direct sensory inputs that bypass the midbrain. The collo- and lemno- receiving areas appear to be evolutionary distinct with unique functions. For example, the extrastriatal visual cortex is a lemno- receiving area, processing visual data from the OT. But because the thalamus also includes areas for the cortex to talk to itself (T.a from the hippocampus to the cortex, T.md for input to the frontal (F) areas, T.re for frontal to hippocampal, T.va and T.vl for internal motor planning), as well as a distinction between direct signal information (“core”) and diffuse context data (“matrix”), it may be best that the model doesn’t try to overcomplicate all those differences.
Frontal and Cortical (“F” and “C”)
The idea between splitting the cortex into frontal and cortical is due to the different connectivity. The frontal areas (F) are more strongly connected with the hypothalamus (H) and its areas, while the other cortical areas (C) either directly control midbrain motor (M) or provide context to areas like the optic tectum (OT).
So, the ventral-medial prefrontal cortex (F.vm) is in the frontal area, as well as the orbital-frontal (F.ofc) and possibly the insular and parts of the cingulate cortex (F.ai, F.i, F.ac), but the dividing lines don’t appear to be particularly clear-cut. The dorsal-lateral PFC (F.dl) seems to be motor based, but “C.dl” makes no sense.
Because it’s the primary olfactory cortex, the piriform is O.pir.
The mnemonic model has been a personal help also in trying to understand the connectivity of regions. As well as grouping similar inputs and outputs (for example areas that project too many hypothalamic nuclei: H.vm, H.dm, H.l, H.pm, etc.), the naming also helps distinguish areas that stand out. For example, areas that come directly from the hindbrain/brainstem like B.pb (parabrachial nucleus) are interesting inputs to areas like the amygdala that mostly receive indirect input. And output areas that project to brainstem areas (B.ret to use the archaic, lazy reticular instead of specific B.xx areas) stand out as actual motor outputs instead of endless internal processing.
References
Bedont, Joseph L., Elizabeth A. Newman, and Seth Blackshaw. “Patterning, specification, and differentiation in the developing hypothalamus.” Wiley Interdisciplinary Reviews: Developmental Biology 4.5 (2015): 445-468.
Butler, Ann. (2008). Evolution of the thalamus: A morphological and functional review. Thalamus & Related Systems. 4. 35 – 58. 10.1017/S1472928808000356.
Kiecker C, Lumsden A. Hedgehog signaling from the ZLI regulates diencephalic regional identity. Nat Neurosci. 2004 Nov;7(11):1242-9. doi: 10.1038/nn1338. Epub 2004 Oct 24. PMID: 15494730.
Puelles, Luis, and John LR Rubenstein. “Expression patterns of homeobox and other putative regulatory genes in the embryonic mouse forebrain suggest a neuromeric organization.” Trends in neurosciences 16.11 (1993): 472-479
Swanson LW. Cerebral hemisphere regulation of motivated behavior. Brain Res. 2000 Dec 15;886(1-2):113-164. doi: 10.1016/s0006-8993(00)02905-x. PMID: 11119693.
Tosches, Maria Antonietta, and Detlev Arendt. “The bilaterian forebrain: an evolutionary chimaera.” Current opinion in neurobiology 23.6 (2013): 1080-1089.