The ascidian circuit in essay 30 had an interesting dopamine subcircuit that looks like an indirect search, where the ascidian coronet cells modulate the underlying phototaxis and geotaxis circuits. While the function of the coronet cells is unknown, if these cells are another seeking system like following an odor, then the coronet sub circuit follows odor by modulating different seek circuits: phototaxis and geotaxis.
Ascidian analogy
Tunicates are the closest non-vertebrate chordates evolutionarily, but they have developed in vastly different directions from the vertebrates, and likely very differently from the shared common ancestor [Holland 2015]. The ascidian tunicates, which are the most studied tunicates, live their asul life as sessile filter feeders like sponges. Their eggs hatch in only 20 hours and their brief tadpole form lasts only for a few hours, just enough to swim and disperse to find a likely permanent settlement place. Their locomotive strategy is to swim up using geotaxis in the morning and swim down using phototaxis in the afternoon. If they’re lucky enough to find a ledge, they swim up into the ledge’s shadow to settle because hanging like a bat from a ledge offers more protection from some predators than resting on the ocean floor [Zega et al 2006].
As would be expected from a 20-hour brain, the navigation circuit is fairly simple. There are two distinct action paths, one for geotaxis using a heavy pigment cell and one for phototaxis using photoreceptors and another pigment cell as a shadow to provide photo-directionality. The two action paths are connected, where dimming produces upward swimming [Bostwick et al 2020].
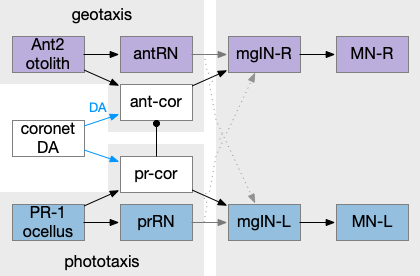
Ascidian tadpole sub circuit for geotaxis and phototaxis. The horizontal neurons are the main action paths. The coronet DA cells modulate the action paths.
In the above diagram, the geotaxis action path starts from the otolith (“ear stone”) receptor ant2, which is functionally similar to the vestibular system (but not related), passes input to antenna relay neurons (antRN) and then to the right side motor neurons (mgIN-R and MN-r) [Ryan et al 2016]. Similarly, the phototaxis action path starts from the ocellus (eyespot) to the phototaxis relay (prRN) and to the left motor neurons, providing an opposing direction from geotaxis. Importantly for the following discussion, each path has a weak connection to the opposite direction, possibly to add some stochasticity to the movement to improve dispersion of the many tadpoles.
The function of the coronet cells is unknown, although they have some genetic connection the palp sensory cells [Cao et al 2019]. Other papers compare the corona cells to dopamine cells in the hypothalamus and Ob (olfactory bulb) [Horie et al 2018] or ancestral photo-hypothalamus and retina [Sharma et al 2019], possibly related to the fish saccus vasculosus area of the hypothalamus, responsible for some circadian behavior. However, the ascidian tadpole has lost circadian clock genes, which argues against circadian timing [Chung et al 2023]. The coronet cells can accumulate serotonin and the DA might promote onset of metamorphosis [Razy-Kraika et al 2012]. So, the coronet may be involved in triggering metamorphic changes at twilight, which causes the tadpole to dive to deeper waters [Lemaire et al 2021].
Whatever the source, the interesting thing about the circuit is that it’s an indirect modulation of underlying taxis action paths. The action of the coronet is gating or modulatory. While this coronet circuit is not homologous to the basal ganglia, using it as an analogy may be useful. For example, dopamine is a sleep / wake signal for the basal ganglia [Vetrivelan et al 2010]. Because low dopamine reduces basal ganglia activity both at the striatum input layer and the Snr (substantia nigra pars reticulata) output layer, it’s an effective sleep controller.
Indirect chemotaxis
Consider indirect chemotaxis, where the animal is seeking toward the odor, but the underlying action path is phototaxis or geotaxis, like the ascidian circuit above. If the animal detects an odor, it increases the current direction. In other words, the current direction is toward or near a food odor. This strategy is like the e. coli tumble-and-run strategy, where the bacteria runs further when the odor gradient is increasing.
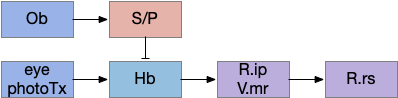
Consider the basal ganglia as an analogy. For example, Ob has some dopamine interneurons (Ob.sac – short axis cells) that project to S.ot (olfactory tubercle) [Burton 2017], a portion of the stratum focused on olfactory input. For the corollary of the phototaxis path, consider the Hb.m (medial habenula) phototaxis path [Zhang et al 2017].
Hypothetical indirect seek circuit where chemotaxis uses an underlying phototaxis to hunt for food. Hb (habenula), Ob (olfactory bulb), P (pallidum), R.ip (interpeduncular nucleus), R.rs (reticulospinal motor neurons), S (striatum), V.mr (median raphe).
When the odor is detected, Ob enables the basal ganglia, which enhances the phototaxis path. If the odor isn’t detected, the default semi-suppressed behavior means the direction is semi-random. This indirect control would allow for seeking odor when the underlying navigation is phototaxis and geotaxis.
Discussion
After writing this description. I think this model may be a bit sketch for something like chemotaxis, although it’s a reasonable model for sleep. Because I’m not sure the idea is likely to be productive, I’m holding off on doing any implementation, but writing down the description in case it makes sense later.
The original impetus for this sleep essay was the idea that the basal ganglia could best be understood as a sleep and wake circuit [Kazmierczak and Nicola 2022]. After reviewing the rest of the brainstem sleep circuitry, it’s time to tackle the original problem.
Snr as a sleep/wake gate
Snr (substantia nigra pars reticulata) is the output node of the basal ganglia. It’s a set of GABA neurons that tonically suppress the majority of all brainstem motor areas including MLR (midbrain locomotor region), OT (optic tectum), and R.rs (hindbrain reticulospinal motor command) with corollary discharge to the thalamus. Snr can inhibit initiation of eating and motion [Rossi et al 2016], but don’t disrupt ongoing actions [Liu et al 2018]. Disruption of Snr can cause hyperactivity and insomnia [Geraschenko et al 2006]. The caudal Snr derives from hindbrain r1 (rhombomere r1 near the midbrain-hindbrain boundary) [Achim et al 2012], [Lahti et al 2015], [Partanen and Achim 2022], suggesting it may be evolutionarily old, possibly older than other basal ganglia regions.
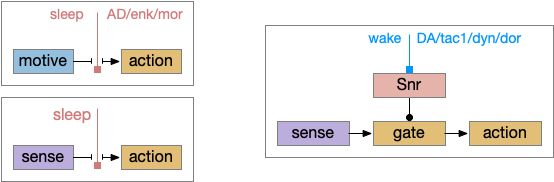
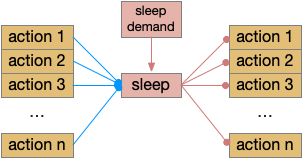
Sleep as gating motive from action or sleep from action. Wake as disinhibiting sleep. Snr (substantia nigra pars reticulata).
As described in part 1 this essay, sleep suppresses senses, motivation and action. To implement this suppression, sleep could disconnect senses and motivation neurons from action neurons. In the above diagram, the gate is conceptual. The circuit could also inhibit the sense or action nodes directly instead of requiring specific gating neurons. This gating architecture has the advantage of simplicity because the sleep circuit can be localized in the gate, while the senses and actions can be mostly free of sleep circuitry.
As a preview, sleep neurotransmitters and peptides in BG (basal ganglia) include AD (adenosine), enk (enkephalin), MOR (μ-opioid receptor), and wake neurotransmitters include DA (dopamine), tac1 (tachykinin 1 aka neurokinin 1 aka substance P), dyn (dynorphin), and DOR (δ-opioid receptor).
If the vertebrate brain follows this architecture, Snr is well-placed to control that gate. Snr.m (medial Snr) projections have many collaterals to distinct motor areas and suppressing the wake-promoting areas covered earlier in this essay, which suggests widespread suppression as opposed to fine-grained control.
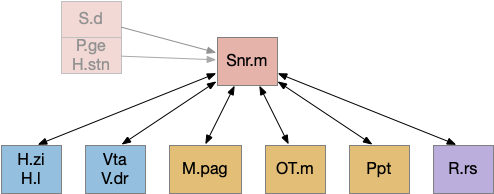
Snr.m gad2 connectivity. 60% of Snr.m inputs are from motor, motivation and wake areas. H.l (lateral habenula), H.stn (subthalamic nucleus), H.zi (zona incerta), M.pag (periaqueductal gray), OT.m (medial optic tectum), P.ge (external global pallidus), Ppt (pedunculopontine nucleus), R.rs (reticulospinal motor command), S.d (dorsal striatum), Snr.m (medial substantia nigra pars reticulata).
As the above diagram illustrates, despite its description as basal ganglia output, 60% of the gad2 (genetic marker), Snr.m inputs are outside of the basal ganglia, particularly from the midbrain (30%) and hypothalamus (10%) [Liu et al 2020]. Snr.m has two independent neuron types marked by gad2 and pv (parvalbumin), which are topographically organized with gad2 in Snr.m and pv in Snr.l (lateral Snr). While Snr.l.pv seems to be strictly motor related, Snr.m.gad2 are sleep related [Liu et al 2020]. However, [Lai et al 2021] reports Snr.l as sleep related.
Functional sleep and action requirements. Any ongoing action should suppress sleep, and sleep should suppress all actions.
Snr’s widespread motor and motivation connectivity suggests a possible primitive role in sleep. Sleep needs to suppress all actions, but any ongoing action needs to suppress sleep, because an animal shouldn’t fall asleep while eating or moving. It seems plausible that a primitive proto-vertebrate could have used Snr for sleep regulation without needing the rest of the basal ganglia.
Because astrocytes can integrate inputs spatially and temporally and are associated with sleep, it’s plausible that Snr astrocyte would be involved in this circuit. Interestingly Snr astrocytes are sensitive to dopamine and become hyperactive in the absence of dopamine [Bosson et al 2015] and are sensitive to glutamate from H.stn [Barat et al 2015].
Dopamine D2.i sleep / wake circuit
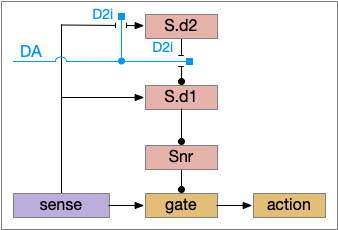
Although the independent Snr circuit is a functional sleep / wake gating circuit, it tonically inhibits the sense to action circuit, adding noise. An improvement to the circuit enables the gate when a signal is available, using the striatum to selectively open the gate. This circuit uses dopamine to open and close the gate. High dopamine is a wake signal and low dopamine is a sleep signal.
In the above diagram, Snr and S.d2 (D2.i associated striatum projection neurons) are sleep-promoting regions and S.d1 (D1.s associated striatum projection neurons) is a wake-promoting region. D2.i (inhibitory Gi-protein dopamine receptor) disconnects inputs, as opposed to inhibiting a neuron directly. When DA is available, S.d2 is disconnected, and S.d1 inhibits Snr, opening the gate. When DA is low, S.d2 is active, which inhibits S.d1, disinhibiting Snr, closing the gate and producing sleep. The D2i between S.d2 and S.d1 is from [Dobbs et al 2016].
The idea of the circuit is that the sense signal disinhibits itself during wake, but sleep prevents sense from disinhibiting itself. The minimal system only requires D2i circuits [Oishi et al 2017]. Wake enables the gate, and sleep disables the gate. Although I’ll cover D1s later, D2i is more fundamental because disabling D1s can be reversed by sufficient arousal, but disabling D2i can’t [Kazmierczak and Nicola 2022].
Note the diagram is somewhat incorrect, because direct S.d2 to S.d1 connection is weak [Tepper 2008]. Instead, S.d2 GABA inhibits S.d1 input at distal dendrites as opposed to inhibiting the neuron soma itself.
P.v ventral pallidum and S.core
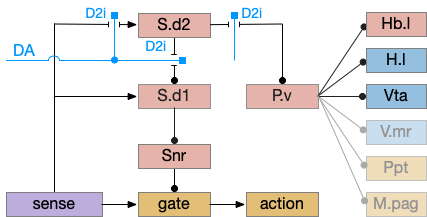
While S.d2 neurons in model above suppresses motor for sleep, S.d2 in S.core (ventral striatum core aka nucleus accumbens) can produce sleep pressure by inhibiting the wake supporting P.v (ventral pallidum) [Oishi et al 2017]. P.v is a tonically active, wake-promoting nucleus, primarily inhibiting sleep areas or disinhibiting wake areas.
Sleep/wake control adding P.v as a tonic wake producing node. DA (dopamine), D2i (inhibitory Gi-coupled dopamine receptor), H.l (lateral hypothalamus), Hb.l (lateral habenula), M.pag (periaqueductal gray), Ppt (pedunculopontine nucleus – ACh), P.v (ventral pallidum), S.d1 (D1-associated striatum projection neuron), S.d2 (D2-associated striatum projection neuron), Snr (substantia nigra pars reticulata), V.mr (median raphe – serotonin), Vta (ventral tegmental area – dopamine).
P.v fill a similar wake-promoting role as S.d1, but unlike S.d1 it’s tonically active and affects the motivation loop of H.l, Hb.l, and Vta instead of gating sense from action. Where P.v supports general wake, S.d1 supports specific wake for an action. Like the previous basal ganglia sub-circuit, this sub-circuit only requires D2i receptors.
P.v promotes wake by inhibiting Hb.l sleep-producing system [Li et al 2023]. It also promotes wake through Vta by disinhibiting GABA interneurons [Li et al 2021]. (It could also disinhibit H.l orexin but I don’t have a reference).
In the model above, stimulating S.d2 inhibits wake-producing P.v, which disinhibits sleep-producing areas like Hb.l and inhibits wake-producing areas like H.l and Vta through GABA interneurons. Conversely, stimulating the D2i receptor by high DA inhibits S.d2, which disinhibits Pv, allowing it so promote wake. Disabling the D2i receptor activates S.d2, promoting sleep even with high dopamine [Qu et al 2010].
Note that S.d1 also connects to P.v and can produce wake [Zhang et al 2023]. P.v has multiple sub-populations with opposing functions. For example, it has both a hedonic hot spot for liked food and a cold spot for disliked food [Castro et al 2015]. For the sake of simplicity the diagram only shows a sleep-promoting path through S.d2, but there may be a wake-promoting path through S.d2 to an opposing P.v subpopulation.
D1s – stimulator dopamine receptors
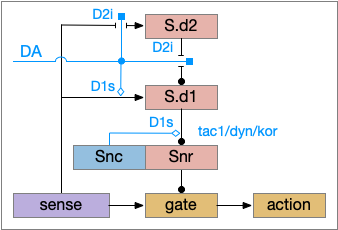
Although using only D2i as a mode switch to the sleep path is functional, it can be improved by also enhancing the wake path with D1s (stimulatory Gs-coupled dopamine receptor).
D1s as enhancing the basal ganglia wake path. DA (dopamine), D1s (stimulatory Gs-coupled dopamine receptor), D2i (inhibitory Gi-coupled dopamine receptor), S.d1 (D1-associated striatum projection neuron), S.d2 (D2-associated striatum projection neuron), Snc (substantia nigra pars compacta – dopamine), Snr (substantia nigra pars reticulata).
The improved circuit works exactly like the D2i-only circuit but enhances the wake path when DA is available. Dopamine boosts both the signals from the sense to S.d1 and the signal from S.d1 to Snr [Salvatore 2024], [Kliem 2007], [Rice and Patel 2015]. When dopamine is available, it boots the sense to S.d1 signal with D1s, which more strongly disinhibits the gate by inhibiting Snr, which is also boosted by D1s.
The D1s in Snr and dopamine may be more important for motor suppression than dopamine in the striatum [Salvatore 2024]. In Parkinson’s disease and also normal aging, bradykinesia (slow movement) correlates with dopamine in Snr more closely than dopamine in the striatum. Motor symptoms in Parkinson’s disease don’t generally occur until striatal dopamine is reduced by 80%, but the effect on Snr is more immediate with only a small drop of dopamine.
Note that the Snc (substantia nigra pars compacta) to Snr dopamine comes from somatodendritic broadcast, not from an axon synapse. Snc dendrites in Snr produce dopamine to enhance the S.d1 to Snr connection.
Although the previous diagrams show the basic logic of the circuit, the basal ganglia use adenosine as a sleep-producing neurotransmitter, competing with dopamine.
Adenosine in striatum sleep
Adenosine is a product of the energy molecule ATP and is produced by neural activity, and also as a astrocyte transmission molecule. Although adenosine can accumulate in a circadian manner, particularly in P.bf (basal forebrain), it’s typically a shorter term sleep pressure. Caffeine is wake promoting by suppressing adenosine receptors.
Dopamine and adenosine are paired, opposing neurotransmitters in the basal ganglia: dopamine produces wake and adenosine promotes sleep. As an opposing signal to dopamine, the adenosine circuit is a flip version of the dopamine circuit.
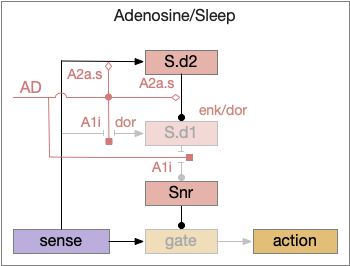
Parallel adenosine sleep circuit in the basal ganglia. AD (adenosine), A1i (inhibitory Gi-coupled adenosine receptor), A2a.s (stimulatory Gs-coupled adenosine receptor), S.d1 (D1-associated striatum projection neuron), S.d2 (D2-associated striatum projection neuron), Snr (substantia nigra pars reticulata).
When adenosine is active in the above circuit, it cuts off S.d1 input and output and enhances S.d2’s suppression of S.d1. With S.d2 fully suppressed, Snr is free to suppress the gate and therefore suppress sleeping action.
Since adenosine is low in the morning, sleep is suppressed, which is enhanced by high ultradian morning dopamine. If A2a.s (stimulating Gs-coupled adenosine receptor) are stimulated in the striatum, the animal is more likely to sleep even in the morning [Yuan et al 2017], specifically in S.core not S.sh (ventral striatum shell aka nucleus accumbens) [Oishi et al 2017].
The dual signal system allows for interesting combinations at the boundary between sleep and wake. If adenosine is high with sleep pressing, then a large amount of dopamine motivation is required to continue wake. In fact, sleep deprivation down regulates D2i receptors, moving from the neuron membrane to the interior [Volkow et al 2012], which tips the balance toward sleep by diminishing the D2i-mediated wake signal. Caffeine inhibits both the A1i (inhibitory Gi-coupled adenosine receptor) and A2a.s receptors, tipping the balance to dopamine wake.
Dorsal striatum indirect path
The full S.d (dorsal striatum) path includes an indirect path, but this path may be more related to pure motor control, not sleep. As mentioned above, Snr divides into two populations Snr.l with pv neurons and Snr.m with gad2 neurons, and the Snr.l neurons are motor related, not sleep related [Liu et al 2020]. Similarly, the indirect path including P.ge (external globus pallidus) and H.stn (sub thalamic nucleus) may not be sleep related. Nevertheless, I’ll include it here, in case it is sleep related.
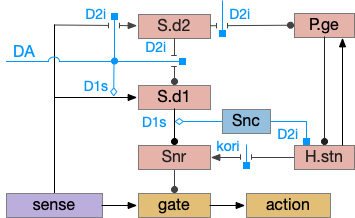
S.d model with indirect path included. DA (dopamine), D1s (stimulatory Gs-coupled dopamine receptor), D2i (inhibitory Gi-coupled dopamine receptor), H.stn (subthalamic nucleus), P.ge (external globus pallidus), S.d1 (D1-associated striatum projection neuron), S.d2 (D2-associated striatum projection neuron), Snc (substantia nigra pars compacta), Snr (substantia nigra pars reticulata).
Note that both P.ge and H.stn are tonically active, and they oscillate together at beta frequencies (roughly 10hz), which suppresses action. An excessive beta oscillation in this P.ge and H.stn circuit is a Parkinson’s disease symptom that suppresses motion and can also interrupt sleep. D2i receptors in H.stn mean that dopamine suppresses H.stn output [Shen et al 2012].
One significant experiment showed that lesioning P.ge increased wake by 40%, particularly eliminating normal circadian night-time sleep, replacing it with day-time like napping [Qiu et al 2016], which would suggest that P.ge is a major sleep center like Po.vl (ventrolateral preoptic area) [Vetrivelan et al 2010]. Note that this analysis would suggest that my basal ganglia sleep diagram is entirely wrong, because P.ge as a sleep center is basically incompatible with its position in the circuit.
P.ge – external globus pallidus
Lesioning P.ge increases wake by 40%, almost entirely eliminating circadian sleep [Qiu et al 2016]. However, this produces hyperactive chewing, weight loss, abnormal motor behavior and death in 3-4 weeks [Vetrivelan et al 2010]. Other manipulations of P.ge produce hyperactivity, abnormal movement, and odd stereotypical behavior [Gittis et al 2014]. So, it’s unclear to me that P.ge is a sleep center, but removing P.ge produces excessive action which then suppresses sleep.
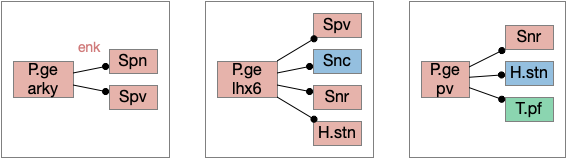
In addition, P.ge is a heterogenous area with at least three major cell types with distinct projections and roles. Arkypallidal neurons project strongly and exclusively to the striatum. Lhx6 neurons project strongly to Snc and to some areas of H.stn, excluding the center. Pv neurons project to all of H.stn and also to T.pf (parafascical thalamus) [Gittis et al 2014].
Distinct projection neuron types of P.ge. H.stn (subthalamic nucleus), P.ge (external globus pallidus), Snc (substantia nigra pars compacta), Snr (substantia nigra pars reticulata), Spn (striatal projection neuron), Spv (pv marked striatum interneuron), T.pf (parafascicular thalamus).
With three projection types, it’s possible that they have entirely separate functions. For example, the lhx6 projections are functionally compatible with a sleep promoting role, and lhx6 neurons in H.zi (zona incerta) are sleep promoting [Liu et al 2017].
Liu K, Kim J, Kim DW, Zhang YS, Bao H, Denaxa M, Lim SA, Kim E, Liu C, Wickersham IR, Pachnis V, Hattar S, Song J, Brown SP, Blackshaw S. Lhx6-positive GABA-releasing neurons of the zona incerta promote sleep. Nature. 2017 Aug 31;548(7669):582-587. doi: 10.1038/nature23663. Epub 2017 Aug 23.