The simple phototaxis implementation exposes a few problem with the simulation, both from running it and from reviewing neuroscience to critique it.
Interrupts
The essay doesn’t currently implement any interrupt mechanism. When running into darkness, the animal turns around and starts an area restricted search (ARS), but if the animal is in the middle of a long Levy path, it will cross the border and the search will not find the border.
Long initial paths breaks the phototaxis algorithm.
The problem here is that the ARS starts too late because the animal doesn’t interrupt the current behavior when encountering the border.
One solution is to create an interrupt (orientation) system, which exists in the vertebrate brain in V.ppt (peduncular pontine nucleus), and uses ACh (acetylcholine) to interrupt the current behavior. A natural location for the interrupt is V.ppt for the signal and the stratum as the plan representation, interruptible via ACh interrupts to the striatum.
Another solution is to avoid the uninterruptible behavior entirely, where the problem is the essay’s Levy walk implementation. The essay pre-computes the length of a run instead of continuously creating extensions. In contrast the zebrafish larva swims in bouts, but longer runs are made of multiple forward bouts.
Zebrafish random walk (ARTR area)
The essay’s random walk does not match actual zebrafish search behavior. The essay uses a turn-and-run model where the turn and run length are computed randomly. The zebrafish has a hindbrain oscillator, the ARTR (anterior rhombencephalic turning region) which selects left and right turns [Karpenko et al 2020].
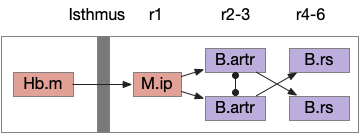
Speculative directed random walk with the zebrafish ARTR. The isthmus is the midbrain-hindbrain boundary (MHB). B.artr anterior rhombencephalic turning region, B.rs reticulospinal motor command, Hb.m medial habenula, M.ip interpeduncular nucleus.
In zebrafish, turns and runs are selected independently and can be chained differently. Instead of turn-run-turn-run as in the essay, the zebrafish can have turn-turn or run-run patters. Zebrafish turn direction is also correlated, as opposed to the random walk’s turn independence. A zebrafish left turn is more likely to follow a left turn.
When encountering darkness, the same-direction turns increase. When encountering light, alternating turns increase. Together with the sharp turn (O-bend) followed b shallower turns, this behavior should create a spiral-like search for the light area.
Note: this specialized circuitry in the hindbrain suggests that random search is a primitive behavior. Although the essay put the Levy walk logic in the midbrain, it belongs in the more primitive hindbrain.
Head direction
[Petrucco et al 2023] report that M.ip is highly connected with head direction axons from B.dtg (dorsal tegmental area of Gudden). This head direction does not receive vestibular input, but is likely derived from motor efferent copies. Since both M.ip and B.dtg are r1-derived regions and possibly ante-vestibular, this head-direction and M.ip connection may be ancient.
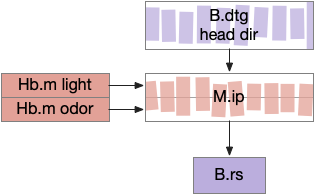
Speculative M.ip circuit following the fruit fly fan-shaped body. B.dtg dorsal tegmental nucleus of Gudde, B.rs reticulospinal motor command, Hb.m medial habenula, M.ip interpeduncular nucleus.
This organization is strikingly similar to the fruit fly’s ellipsoid body (EB), protocerebral bridge (PB), and fan-shaped body (FB) in the central complex (CX) [Hulse et al 2021]. EB and PB calculate head direction. The fan-shaped body merges head direction with goal direction to produce motor commands. In this diagram, M.ip represented as if it resembles the fan-shaped body.
If the M.ip functionality is similar to the fan-shaped body, it’s highly likely to be convergent evolution, not homology because amphioxus lacks any similar structure.
Dark search
When zebrafish are plunged into darkness, they initiate a search that continues for about five minutes. The darkness behavior increases speed and straight behavior [Horstick et al 2017]. In other words, phototaxis is not just gradient behavior but also has steady-state darkness behavior. Because zebrafish require light to hunt, darkness in itself is is an area to avoid.
The essay is purely gradient based and has no speed changes. Photokinesis is moving faster in darkness and slower in light, which will bias the time spent in the light area.
Previous essays used a simple default ballistic forward motion action: the animal moved forward until it hit an obstacle or encountered food or an odor plume. Furthermore, all actions in the essay were continuous: at every time-step direction or speed could change. The animal was slug-like. However, since vertebrate is oscillatory — swimming or walking — powered by central pattern generators (CPGs), vertebrate motor commands are not continuous but modifiable only on correct timing of the oscillatory cycle.
This essay 21 adds some more realism to the simulated animal by creating distinct action syllables, each of which runs to completion. In addition, the default movement is a Lévy walk to more efficiently search for food.
Zebrafish bouts and mouse modules
Zebrafish larva move in discrete bouts [Johnson et al. 2020], punctuated by pauses. Each bout is on the order of 200ms to 1000ms and consists of stereotyped movement like a forward swimming stroke, and several turn types possibly followed by forward motion. The neural source of the bout timing is not know, although basal ganglia (striatal) defects can also produce jerky motion instead of smoothly linked motion.
Mouse movement is also comprised of small modules [Wiltschko et al. 2016] on the order of 60 modules in open exploration, each module lasting 200ms to 500ms. Unlike the zebrafish larva, sequences of mouse modules are linked smoothly instead of in jerky bouts.
Simulation action syllables
To match the vertebrate action syllables, the essay simulation now has action syllables at the lowest layers. The action syllables are fixed programs that last for several simulation ticks. For example a forward left turn might take 10 simulation ticks, depending on the tick resolution. In other words, these syllables are more like real-time actions with limited time resolution, not turn-based actions like a board game.
Unlike the current action completes, the system ignores new neural commands. In the future, specialized commands like freeze or panic escape might override the current action, like fast escape of zebrafish that bypass normal motor logic. In theory the system could also incorporate mid-action modulation, such as adding power to a swimming stroke without altering the basic action.
The new syllables should add realism and also introduce complications that vertebrates need to solve, such as timing for sequences of syllables. The syllables also introduce issues because prior-action memory as modeled by the striatum and nucleus isthmi need to persist for a syllable, not just a simulation tick.
Lévy walks
Along with the syllable changes, essay 21 adds better default movement. Previously if the animal wasn’t approaching food or avoiding an obstacle, it would move forward ballistically. That simplification only worked because the simulation is a simple bounded box, but in nature animals have better default search strategies.
Brownian motion is a simple default. The animal turns a random direction, then moves forward a random (gaussian) distance. Brownian motion does well at searching a neighborhood and is better than the essay’s ballistic strategy, but it tends to get stuck in a small area. When food is scarce and patchy, the brownian strategy won’t move the long distances needed to effectively find a new patch. The Lévy walk improves on this strategy by moving long distances when the current neighborhood is already searched [Abe 2020].
Default random walk for the simulated animal.
A Lévy walk is fractal (“scale-free”): larger walks have the same search structure as neighborhood search. If described in probabilistic terms, the distance traveled looks like a power law:
P(len) = 1/ len ^ 2
Where the exponent 2 (alpha) can be generalized between 1 and 3. The bounds are because the exponent 1 or less is ballistic and exponent 3 or more is essentially brownian. If alpha is closer to 1, the search is wider and moving further, while an alpha is closer to 3, the search is closer and more local.
Central pattern generators, criticality, and chaos
Animals do seem to use Lévy walks [Kölzsch et al. 2015] and the source appears to be internally generated [Berni et al. 2012] as opposed to a response to a fractal environment. The internal source of the randomness is not well know, although central pattern generators appear to be a possibility [Sims et al. 2018], [Reynolds 2019].
Near a critical point, fractal patterns appear and can be efficient computationally [Abe 2020], and a relatively simple chaotic system with only two random variables and produce Lévy walks. More broadly, other brain areas such as the cortex may also use criticality or near criticality to improve computation and generate longer-lasting signals to solve the timing problem [Hidalgo et al. 2014]. The timing problem is that fast neurons are 10ms but behavior needs to be responsive on the order of seconds and minutes.
Vertebrate source of Lévy walks
Although there is some knowledge of the search circuitry in fruit flies [Berni et al. 2012], the vertebrate circuitry seems entirely unknown.
As a thought experiment, consider the midbrain theta circuitry as part of the exploration circuit [Cisek 2022], and therefore related to the Lévy walk. If the vertebrate search is generated by a combination of central pattern generators then the source should be in the hindbrain, near those CPGs.
Theta clock cycles (4-12hz) in the brain are strongly correlated with exploratory movement, thought, and learning especially with the hippocampus (E.hc).
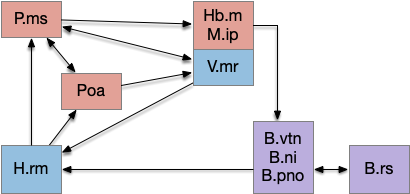
In the above, the hindbrain contains the chaotic search circuitry and generates theta, such as B.pno and B.vtn in conjunction with the reticulospinal motor command (B.rs). B.vtn is the forward movement analogue of the head direction nucleus B.dtn.
The medial septum (P.ms) is essentially the main theta clock for the hippocampus. Part of the theta cycle is generated internally to P.ms with spontaneous interaction of acetylcholine (ACh) and GABA inhibitory neurons, but exploration-related theta comes from the hypothalamic retromammilary (H.rm, aka supramammilary), which receives its theta from the hindbrain theta centers.
The movement restriction for theta depends on the median raphe (V.mr), which is one of the two main serotonin (5HT) centers. If V.mr is disabled, theta through H.rm always exists, not restricted to exploration. V.mr is in turn strongly influenced with the habenula (Hb.m) and interpeduncular (M.ip) complex.
Simulation
For now, I’m avoiding using criticality or chaotic variables in the simulation, although criticality is an interesting design to explore and very possibly how the vertebrate brain solves these problems.
The disadvantage is that the simulation would quickly become unclear and overcomplicated. While a few chaotic variables in the brainstem might be manageable, extending that idea to the striatum and cortex seems like it would become impenetrable. Since the purpose of the simulation is something like an executable thought experiment or executable diagram, and impenetrable simulation is defeating the purpose. Some alternatives like [Bartumeus and Levin 2008] fractal reorientation clocks might serve the purpose while remaining more clear.