The model required a major refactoring to properly simulate the essay.
Model update to simulate essay 27.
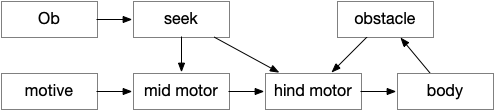
HindMotor
HindMotor includes the motor command areas in the hind-brain with locomotion and eating as separate modules. Locomotion models B.rs (reticulospinal motor command neurons) and B.mdd (medulla reticular neurons). Eating models parts of B.pb (parabrachial nucleus), B.nts (nucleus of the solitary tract) and B.mdd. The locomotion and eating modules do not coordinate at the hind-brain level for this model.
In this essay, HindMotor controls the random search, modulated by upstream request, and also manages action bouts. One an action starts, it continues until complete, which generally requires several simulation ticks. Because the essay model is real-time, not turn-based, actions require several simulation ticks to complete.
HindMotor locomotion commands are split between directional hints and forward movement hints, following a similar division in vertebrates. Turn modulation comes from target seeking and obstacle avoidance, either encouraging or inhibiting left vs right turns. Forward movement modulation comes from the motive core, specifically selecting between a roaming search or an area-restricted dwelling search.
MidMotor
MidMotor coordinates actions that HindMotor implements. MidMotor with sustains actions across action bouts, and manages the transition between action bouts. Because ongoing movement needs to stop before eating, MidMotor pauses eating until the animal stops.
MidMotor represents Ppt (predunculopontine nucleus), H.stn (subthalamic nucleus), OT.d (deep optic tectum), MLR (midbrain locomotor region), and T.pf (parafascicular nucleus). For this essay, Ppt and H.stn work together as a single module to sustain actions and pause upcoming actions until currently-active actions are complete.
Seek
The essay Seek is directional movement toward a specific target, the same idea as taxis (but avoiding Greek). Seek is only active with a specific directional cue, here an olfactory gradient.
Seek models Hb.m (medial habenula) and M.ip (interpeduncular nucleus), where M.ip is models as a gradient seek module like the Drosophila fan-shaped body.
CoreMotive
CoreMotive simulates the motivational core, which is primarily peptide based. In this essay, the neural areas include H.l (lateral hypothalamus), V.dr (dorsal raphe), and B.pb (parabrachial nucleus). H.l is strongly associated with all aspects of feeding and is the driving controller. V.dr expresses the dwell state, which restricts search to a small area once the animal has found food. B.pb manages eating, taste, and physical alarms that might interrupt eating.
Motive neuropeptides
Because the motive core is more broadcast neuropeptide-based than a connective circuit, the simulation includes broadcast neuropeptides as primitive motives. In this essay, the key motives are Roam (motivation to search for food, orexin), Dwell (area restricted search, serotonin), Seek (tracking a target, dopamine) and Sated (antagonizing all food search, GLP-1).
Each motive is a DecayValue, which represents a slow leaky integrator, where the decay time can be tens of seconds or longer, because neuropeptide timing can be long. To a Dwell signal might last for 20 seconds or more without requiring recurrent neural behavior to maintain the state. Since these Motives are broadcast, they can modulate any module with requiring a direct connection.
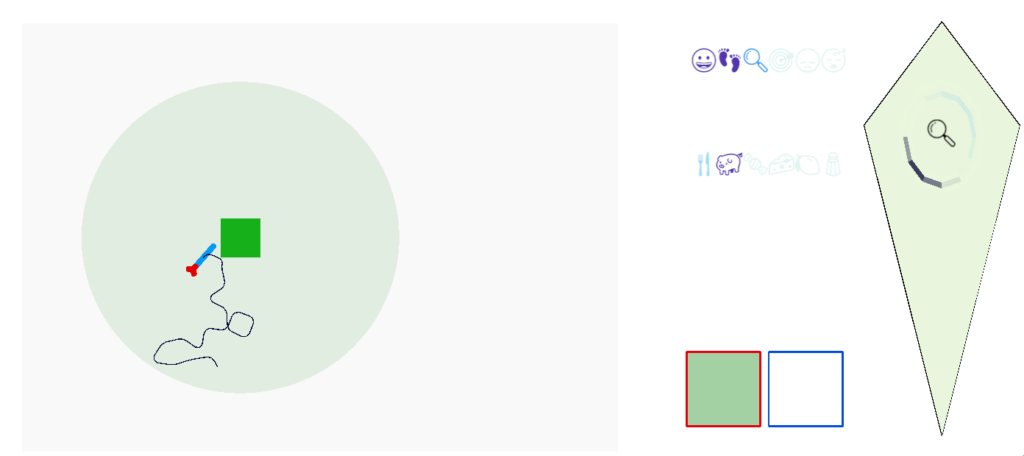
Screenshot after eating, showing multiple active motives.
The screenshot above shows several motives after the animal eats, where emojis represent the active motives. The animal is sated (pid), eating is fading (faded fork and knife), search is in dwell (magnifying glass) because the animal has just eaten, and it’s still roaming (footprints).
Essay 27 returns to feeding, which essay 23 had an earlier sketch of. While the animal in earlier essays could eat while moving, like snails and worms, this essay will add the requirement of stopping before eating, which requires extra control mechanisms to manage the state transition.
A filter feeder like amphioxus, a non-vertebrate chordate that may hint at pre-vertebrate feeding, might move to find a better feeding zone, but then settles down as a static filter feeder. Tunicates, which are more closely related to vertebrates settle down permanently as adults and dissolve their brain as no longer needed. Because I want to keep the essay simple, I’m imaging something more like licking, which is more studied in rodents, as opposed to a more alien filter feeding. The main problem for the essay to introduce locomotion and eating as distinct actions.
As a contrast to further explore the idea of states and state transitions, the essays also explores the transition between roaming and dwelling: global wide-ranging search vs area restricted search. Roaming and dwelling are more amorphous motivational states as opposed to the strict motor division between moving and eating.
Feeding states
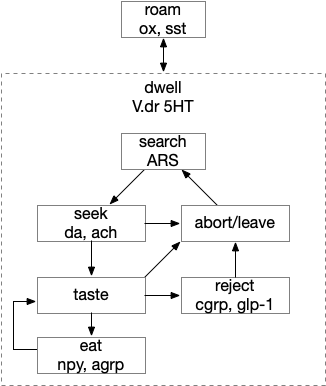
Below is a more detailed diagram of the foraging and feeding states, revolving around the core foraging task. The animal passively roams until is finds an odor cue for a food target, which starts a seek to the target. If it finds food, the animal sops and eats.
In this model, the roam state and dwell state can be separate from seeking a target, depending on the animal’s environmental niche. A seek can start in a roam state or a dwell state, and seek cues may or may not initiate dwell state. For example, dwell state might only start when the animal eats nutritious food, indicating that food is nearby.
Feeding state diagram for the essay. ach (acetylcholine neurotransmitter) agrp (hunger peptide), ARS (area restricted search), cgrp (alarm/bitter taste peptide), da (dopamine), glp-1 (satiety/sickness peptide), ox (orexin wakefulness/action peptide), set (somatostatin peptide), V.dr (dorsal raphe), 5HT (serotonin)
The diagram includes important failure states. If seeking fails, the animal gives up and leaves the area, and must ignore the last cue to avoid perseveration. If the taste is bitter or toxic, the animal rejects the food. For now, I’m postponing longer failure states like the food lacking nutritional value or causing food poisoning.
To avoid perseveration, seeking the failed cue forever, the avoid state moves the animal away from the failed cue and ignores seek cues. A more sophisticated brain could remember the failed cue for a short time, but the current essays lack short term memory.
Eating here means specifically licking or filter feeding. I’m being precise here because the simulation requires it, and more vague neuroscience terms like “reward” are often unclear about exactly what it’s relation to actual eating are.
The connection between the dwell state and serotonin is from [Flavell et al 2013], [Ji et al 2021] which founds serotonin marking the dwell state in the flatworm C. elegans, and [Marques et al 2020] finding serotonin for a zebrafish dwell (“exploit”) state.
Roaming and dwelling
Food search phases have multiple strategies, broadly divided into roaming and dwelling. Roaming is a broader, more general search without a specific area or target. Dwelling or ARS (area restricted search) is slower, with tighter turning, where the current area is believed to be more likely to have food. [Horstick et al 2017] describes dwell as four properties: reduction in travel distance, increased change in orientation, increased path complexity, and a directional bias.
For this essay, dwelling is a motivational drive not a motor command, meaning it can overlap with other motivations and doesn’t provide a strict action state requirement. For example, dwell isn’t required to seek a target, which can occur in the roaming state, for example in C. elegans [Ji et al 2021].
In the C. elegans the dwell state is associated with serotonin and the roam state with PDF (pigment dispensing factor) [Flavell et al 2013]. In zebrafish the dwell state is associated with V.dr (dorsal raphe) serotonin [Marques et al 2020], the roam state is associated with SST (somatostatin peptide) [Horstick et al 2017]. While arousal isn’t quite the same as well, [Lovett-Barron et al 2017] found SST as a low-arousal marker, while CART, ACh (acetylcholine), NE (norepinephrine), serotonin, dopamine and NPY (neuropeptide Y) as signs of high arousal.
Triggers for the dwell state depend on the animal’s species [Dorfman et al 2020]. In C. elegans, which feeds on bacteria, nutritional feedback extends the dwell state [Ben Arous et al 2009]. In some animals a food cue triggers dwell, while in others only eating nutritious food triggers dwell. In zebrafish lack of a food cue causes H.c (caudal hypothalamus) activation decay [Wee et al 2019].
Reflexive eating
This essay models reflexive eating as a hindbrain system controlled by B.pb (parabrachial nucleus) with downstream motor and sensory in B.nts (nuclei tractus solitarius), M.mdd (reticular medulla), and B.3g (trigeminal – orofacial sensorimotor). The simulation isn’t as detailed, treating the hindbrain eating as a single low-level module.
This innate circuit can with without input from higher areas [Watts et al 2022]. For example if rodents lack any dopamine, they won’t move or eat and will starve even if food is near them. However, if food or water is placed at their lips, which activates the innate circuit, the rodents will eat [Rossi et al 2016].
The B.pb area also processes sweet, bitter or salt, and can reject food without requiring higher areas. The higher areas modulate B.pb behavior, such as suppressing B.pb’s innate rejection of sour when drinking lemonade.
Because the B.pb innate eating and the MLR (midbrain locomotor region) are independent, some system much coordinate switching between moving and eating.
The illusion of state machine atomicity
The feeding state diagram suggests a simple atomic transition from seeking food to eating the food, but this transition needs management from some neural circuits. For example, when braking during driving, drivers need to pay attention to the stopping distance. Braking stops a car, but the state transition isn’t a simple atomic transition. For this essay’s eating task, some neural circuit must keep track of the animal’s stopping after seeking and only allow eating when locomotion has stopped.
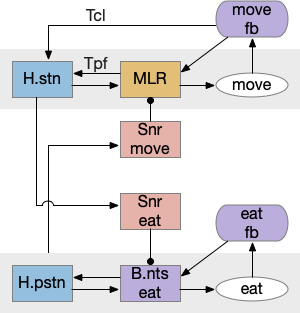
State transition from seeking to eating, emphasizing the stopping state. H.pstn (parasubthalamic nucleus), H.stn (subthalamic nucleus).
H.stn (subthalamic nucleus) is involved with stopping, waiting, and switching tasks [Isoda and Hikosaka 2008]. Since H.stn also receives motor efference copies via T.pf (thalamus parafascicular nucleus) and Ppt (peduncular pontine nucleus), H.stn is in a good position to manage the stopping transition and can prevent eating until the locomotion has ended. The diagrams shows H.pstn (parasubthalamic nucleus) as a parallel area for gaiting eating, following [Barbier et al 2021].
H.stn and H.pstn state transition circuit
H.stn and H.pstn are well-placed to fulfill the transitions between seeking and eating. To flesh this idea out, here’s a simplified model of the seal to eat state transition circuit.
The main action paths are horizontal: moving is from H.stn to MLR to B.rs (reticulospinal motor neurons) and eating is from H.pstn to B.nts to orofacial licking motor neurons. The rest of the circuit manages the transition between the two states.
State transition circuit for move state to eat state. B.nts (nucleus tractus solidarus), fb (feedback), H.pstn (parasubthalamic nucleus), H.stn (subthalamic nucleus), MLR (midbrain locomotor region), Snr (substantia nigra pars reticulata), T.cl (centrolateral thalamus), T.pf (parafascicular thalamus).
Control over the transition comes from S.nr (substantia nigra pars reticulata), which inhibits eating when the animal is moving, and inhibits moving while the animal is eating. To know when the animal has stopped moving, H.stn receives motor efferent copies from T.cl and T.pf (centrolateral and parafascicular thalamus, aka intralaminar). As a note, T.cl contains cerebellum output, so H.stn may receive fine-grained motor timing feedback. H.pstn receives parallel eating efferent copies from B.pb and B.nts to know when the animal has stopped eating.
This circuit has the same structure as a lateral inhibition decision circuit, but the function is about handling timing and transition, not deciding between competing options.
Note: [Shah et al 2022] suggest H.pstn is more specific to suppressing feeding for aversive situations like food poisoning or a predator threat, but not the motor control as described here.
A note on this model: the actual neural circuit isn’t as clean, parallel and logical, because evolution isn’t an intelligent designer. Furthermore, this brain region is part of the neuropeptide core, where neuropeptide broadcast-like signaling can be more important than point-to-point circuit diagrams. Specifically, the disinhibition of B.pb eating is more likely peptides from the hypothalamus, not S.nr tonic inhibition.
H.l food zone
Studies on H.l (lateral hypothalamus) show two interesting results relevant here [Jennings et al 2015]:
Two distinct GABA neuron populations gate eating and seeking.
Two distinct neuron populations are active in a food zone or outside a food zone.
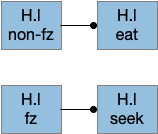
The food zone neurons partially explain how H.l decides between seeking and eating. How does this animal knows when it’s reached the food? In C. elegans there are dopamine chemosensory neurons that sense when the animal passes over food bacteria, and signals the animal to slow [Sawin et al 2000]. Dopamine chemosensory neurons also signal for the animal to turn more when leaving food (dwell-like state) [Hills et al 2004]. For this essay, using B.pb and B.nts to sense nearby food seems like a reasonable simplification because the simulation animal is aquatic and aquatic taste is a chemosensory system, similar to a close-range olfaction.
Food zone modulation of seeking and eating. fz (food zone), H.l (lateral hypothalamus).
The essay uses a signal when the animal is in a food zone or not in a food zone. The food zone signal inhibits eating or seeking actions when the animal is in a non-appropriate place. The essay uses a signal from B.pb as mentioned above.
In mammals H.l receives input from more sophisticated location systems than a bare chemosensory signal, such as E.sub.d (dorsal subiculum of hippocampus), S.ls (lateral septum, which processes hippocampal output), A.bl (basolateral amygdala, highly connected to hippocampus), S.msh (medial shell striatum receiving large hippocampus input) as well as the bare B.pb as for the simulation. All these areas incorporate more complicated environmental context. When the essays start investigating environmental context, I’ll need to revisit the H.l food zone with more sophisticated input.
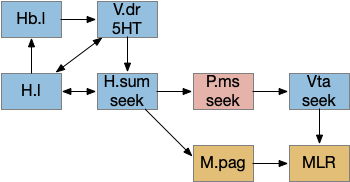
H.sum as driving seek
Fleshing out the drivers of the seek circuit, consider H.sum (supramammillary nucleus, aka retromammillary) and its role in exploring (roaming and seeking). [Ferrell et al 2021] study a subset of H.sum neurons that express tac1 peptide (tachykinin, aka substance-P or neurokinin). These H.sum neurons correlate highly with movement velocity, a second before the action. Since they precede action, they’re upstream in the locomotive path.
H.sum also participates in threat avoidance [Escobedo et al 2023], but that circuit is through Poa (preoptic area) and is outside this essay, although it would be interesting if any of the downstream circuitry is shared. H.sum is also well know for its role in hippocampal theta oscillations, novelty [Chen et al 2020], temporal and spatial memory [Cui et al 2013], and social memory, although those are outside the scope of this essay.
The diagram below shows a possible explore-related path of mammalian H.sum via the tac1 neurons.
It may be important that H.sum and Vta (ventral tegmental area) are both neighbors and H.sum includes dopamine neurons and those dopamine neurons are sometimes considered an extension of the Vta [Yetnikoff et al 2014].
The following diagram gives an extremely rough idea of the adjacency of these areas. In a smaller primitive pre-vertebrate, these might not only be neighbors but mingled earlier functionality. The diagram includes H.zi (zona incerta) because it’s a neighbor, and also because H.zi is a food-seeking area [Ye et al 2023], but I’m postponing consideration of H.zi to a future essay.
Neighbors of the lateral habenula and supramammillary nucleus. H.l (lateral hypothalamus), H.pstn (parasubthalamic nucleus), H.stn (subthalamic nucleus), H.sum (supramammillary nucleus), H.zi (zona incerta), MLR (midbrain locomotive region), Ppt (Pedunculopontine pontine nucleus), Snc (substantia nigra pars compacta – dopamine), Snr (substantia nigra pars reticulata), Vta (ventral tegmental nucleus – dopamine), ZLI (zona limitans intrathalamica).
In addition, the rostral part of Vta nearest H.sum is part of p3 in the prosomeric embryonic model, which is a source of hypothalamic cells [Kim et al 2022]. For pre-vertebrates in this essay, then, there might not be a distinct between H.sum and Vta / posterior tuberculum, particularly since the essays are currently focusing on downstream connections, not upstream dopamine to a future striatum. Zebrafish downstream dopamine circuits directly modulate locomotor movement [Ryczko et al 2020], [Reinig et al 2017]. I think it’s reasonable to simplify this circuit for now and consider H.sum as directly projecting to MLR.
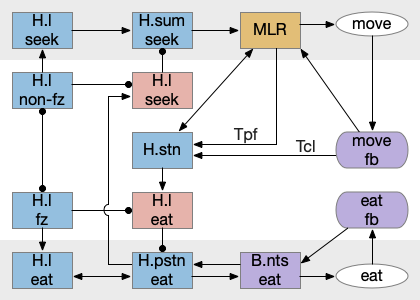
State transition circuit for seek to eat
Putting these ideas together yields something like the diagram below. Like the earlier simplified diagram, horizontal paths drive core seeking and eating behavior, and other circuits manage the state transition. Seeking uses the top path from H.l to H.sum to MLR to B.rs, which produces the final locomotion. Eating uses the bottom path from H.l to H.pstn to B.nts, which controls reflexive eating.
The left contains motivational drivers. The food zone and non food zone systems restrict seeking and eating, only allowing seeking and eating in appropriate locations.
In the center H.stn and its parallel H.stn enforce a smooth transition between seeking and eating, using motor efferent copies to pause transition until active motor stops. The smooth transition creates the illusion of an atomic state transition.
As a diagram note, I’ve used red for the H.l inhibitory neurons that gate seek and eat because they’re playing the same role as Snr neurons. Technically they should be blue, if following normal essay conventions.
Modulation of eating
The eating and feeding modulation systems are complicated and overlapping, which is too detailed for this essay, but two part are interesting. First, B.pb tonically inhibits eating with the CGRP peptide to B.nts. To enable eating, H.arc (hypothalamus arcuate) disinhibits B.nts eating by sending AgRP (a hunger peptide) to B.pb [Campos et al 2016].
Modulation of reflexive eating. AgRP (a hunger peptide), B.nts (nucleus of the solitary tract), B.pb (parabrachial nucleus), CGRP (an anti-eating peptide), H.arc (hypothalamus arcuate).
Although the essays have used the disinhibition pattern before, the pattern has generally ben GABA disinhibition, while this feeding disinhibition uses peptide signaling. As mentioned above, there are many feeding-related peptides that inhibit, excite, and modulate the feeding system without using connection based synapses.
As a parallel, a drinking modulation path goes through the basal ganglia Snr and OT (optic tectum) [Rossi et al 2016]. This path though the basal ganglia and OT coordinates anticipatory licking, while the earlier B.nts path is reflexive eating.
Control of anticipatory licking. B.mdd (medulla licking motor), OT.dl (deep, lateral optic tectum), Snr.l (lateral substantia nigra pars reticulata)
Another drinking path involves S.a (central/striatal amygdala), midbrain, and hindbrain circuits [Zheng et al 2022]. M.dp (deep mesencephalic nucleus) extends licking but doesn’t initiate it. So M.dp might extend eating after tasting. Similarly B.plc extends eating [Gong et al 2020]. S.a sst (somatostatin peptide) neurons promote eating and drinking [Kim et al 2017].
Sustained eating with an amygdala circuit. B.mdd (medulla motor eating), B.pb (parabrachial nucleus), M.dp (deep mesencephalic nucleus), S.a.sst (set-expressing neurons of the central amygdala).
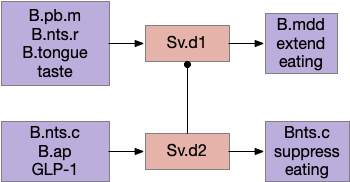
Another path for tasting and eating runs through S.v (ventral striatum). [Sandoval-Rodríguez et al 2023] founds S.v directly controlling feeding using hindbrain taste input to extend eating, and using hindbrain GLP-1 (anti-eating peptide) to inhibit eating. Unlike most striatum circuits, these striatum neurons project directly to the hindbrain motor areas.
Ventral striatum taste exciting and food inhibition circuit with the hindbrain. B.ap (area postrema – nutrient sensing), B.mdd (medulla motor), B.nts (nucleus of the solitary tract), B.pb (parabrachial nucleus), Sv (ventral striatum / nucleus accumbens).
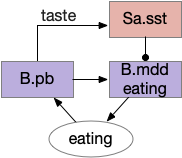
Because this essay is already complicated enough, this simulation isn’t covering all of these details. For simplicity, the simulation will use a simple continuation circuit inspired by the central amygdala and postpone other control circuits for later exploration.
Simplified eating continuation circuit with the central amygdala. B.mdd (medulla motor), B.pb (parabrachial nucleus), Sa.sst (central amygdala, sst projecting neurons)
The important point for now is that eating modulation uses multiple paths, some controlled through synaptic circuits and others through broadcast motivational peptides. The system is not one or the other, but a messy combination. To model this messiness, the simulation needs to handle both systems.
Chen S, He L, Huang AJY, Boehringer R, Robert V, Wintzer ME, Polygalov D, Weitemier AZ, Tao Y, Gu M, Middleton SJ, Namiki K, Hama H, Therreau L, Chevaleyre V, Hioki H, Miyawaki A, Piskorowski RA, McHugh TJ. A hypothalamic novelty signal modulates hippocampal memory. Nature. 2020
Farrell JS, Lovett-Barron M, Klein PM, Sparks FT, Gschwind T, Ortiz AL, Ahanonu B, Bradbury S, Terada S, Oijala M, Hwaun E, Dudok B, Szabo G, Schnitzer MJ, Deisseroth K, Losonczy A, Soltesz I. Supramammillary regulation of locomotion and hippocampal activity. Science. 2021 Dec 17;374(6574):1492-1496.
I like the model of motivation where the hypothalamus and midbrain structures like the ventral tegmental area (Vta), periaqueductal gray (M.pag), and parabrachial nucleus (B.pb) form the motivational core, primarily run by neuropeptides signaling, based on old chemical communication. [Damasio and Carvalho 2013] consider this area as an organized map for feelings, like the optic tectum (OT) has a retina-centric map for visual interest.
To avoid getting stuck in philosophical woo, I’m avoiding the question of whether this area is a primary source of feelings, but I like the idea of a semi-organized map at the base of motivation. The parabrachial nucleus (B.pb) is a good place to start, because its neurons encode warnings like pain, visceral summaries, and primitive feeding, including basic taste.
Parabrachial nucleus
B.pb provides a coarse summary of taste, pain, temperature, and visceral feelings like malaise without the details. It can report that something tastes good because it’s sweet or tastes bad because it’s bitter, but can’t experience chocolate. It’s more of an action-focused alarm [Campos et al. 2018] than a sensory experience.
For example, if B.pb detects bitter taste or malaise, it sends a general notice to other areas in the peptide core to stop eating and investigate further. If B.pb tastes sweet, it encourages eating. In addition to senses like taste and warning, B.pb has action control of its own, including reflexive escape actions, breathing and heart rate to the medulla (B.mdd) and B.nts. So, it can serve as a lower-level action hub.
B.pb and neuropeptides
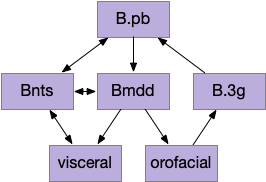
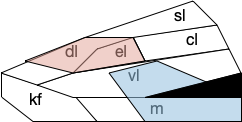
B.pb poses an immediately difficulty for the simulation animal because it’s organized chemically by neuropeptides instead of a simple topological and connectivity map. The following diagram is a broad topographic map of B.pb [Chiang et al. 2019] that illustrates the issue.
Topological map of the parabrachial nucleus.
As shown above, the colored areas do not respect the named boundaries. The blue area represents taste neuron areas and the red area represents general alarm (pain, heat, cold, malaise, etc.) But even those colored areas are an oversimplification because neuron functions are mixed together salt-and-pepper style. [Pauli et al. 2022] found 21 subclusters of B.pb neuron peptide receptors and transmission, each of which may have distinct projection patterns.
This neuropeptide focus isn’t restricted to B.pb. The lateral hypothalamus (H.l), another major node in the feeding circuit, is also organized by neuropeptides, including important ones like orexin (exploring), and MCH, which it sends across the entire brain. Although [Diaz et al. 2023] has broken H.l into 9 areas, these may not be sufficient because of the neuropeptide focus. [Mickelsen et al. 2019] found 15 clusters of glutamate neurons and 15 clusters of GABA neurons. [Guillaumin and Burdakov 2017] and [Burdakov and Karnani 2020] find H.l functional communication through neuropeptides that are invisible to traditional synaptic communication.
Neuropeptide core
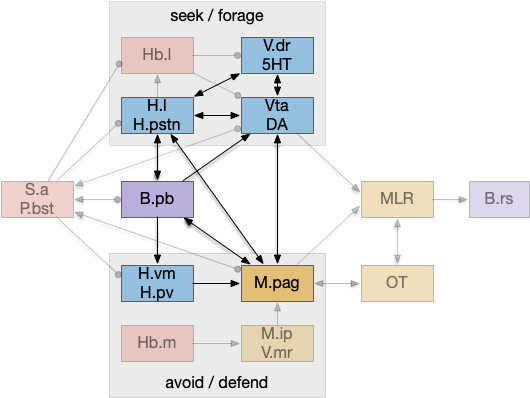
An “isodendritical core” [Ramón-Moliner and Nauta 1966] in the hypothalamus and midbrain is an old idea with a more modern description in [Agnati et al. 2010], which is a good starting point for the essay simulation. The core includes reticular areas of the hypothalamus, B.pb, M.pag, and the Vta (aka posterior tuberculum in zebrafish). “Neuropeptide core” matches my imagination of this area better than the old name. A diagram of the core is below, with the caveat that neuropeptide broadcasting is more important for communication than the diagram’s arrows.
Neuropeptide core in bright colors, associated areas greyed out. B.pb parabrachial nucleus, B.rs reticulospinal motor command, H.l lateral hypothalamus, H.pstn parasubthalamic nucleus, H.pv periventricular nucleus, H.vm ventromedial hypothalamus, Hb.l lateral habenula, Hb.m medial habenula, M.ip interpeduncular nucleus, M.pag periaqueductal, MLR midbrain locomotor region, OT optic tectum, P.bst bed nucleus of the stria terminals, S.a central amygdala, V.dr dorsal raphe (serotonin), V.mr medial raphe, Vta ventral tegmental nucleus (dopamine).
As shown above, the neuropeptide core is highly interconnected. B.pb includes taste and visceral sensation like nausea together with visceral control. H.l includes blood sensors like glucose level, insulin, and fat and protein levels. M.pag includes many innate behaviors including freezing, flight and grooming. Vta controls actions, including seeking and searching.
As the diagram illustrates, the neural connectivity of the inner core is not particularly useful because they’re all entirely interconnected. For simplicity of the essay simulation, I’m using a model where the core neuropeptides are shared in a common neuropeptide soup, or canal, where the neuropeptide identity is more important than the neuron’s specific physical location. For example, treating B.pb as one or two areas instead of the seven areas above.
Cerebrospinal fluid as neuropeptide canal
The periventricular areas like H.pv and M.pag are named for their location around the ventricles, which contains cerebrospinal fluid (CSF). These areas contain neurons that directly sense neurotransmitters and neuropeptides in the CSF itself. The CSF can be a canal for transmitting neuropeptides [Bjorefeldt et al. 2018].
Earlier photo-vertebrate animals may have used a similar canal more extensively. Because of the smaller brain size, diffusion in the canal may have been sufficient for communication without point to point synapses. [Vigh et al. 2004] point out that amphioxus larva, a pre-vertebrate chordate, has much of its com munition in a single neuropile (intertwined dendrites and axons) that’s open to sea water until its neural tube closes as an adult.
Neuropeptides and timing
Neuropeptides act on a much slower timescale than faster neurotransmitters like glutamate and GABA. Glutamate and GABA synapse are a few microseconds and clear rapidly. Neuropeptides can persist tens of seconds to tens of minutes. For an animal’s motivation, like fleeing a predator, the longer timescale is more appropriate, because the animal shouldn’t stop fleeing if it loses sight of the predator for ten milliseconds or even a second or two. Similarly, foraging for food is a longer task measured in many minutes or hours, not milliseconds. The longer chemical timing of the peptides is more suited to motivational timing than the fast reactive transmitters.
Peptide circuits
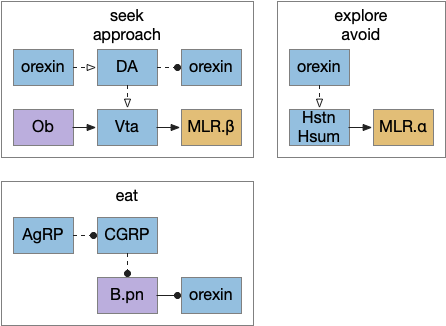
I’ve sketched out some possible neuropeptide circuits for feeding are portrayed in the diagram below, organized by behavior.
Sketch of some of the neuropeptide circuits related to feeding.
The first diagram shows dopamine as a primary seeking neurotransmitter [Alcaro et al. 2007]. When the animal finds target by odor in the simulation, dopamine tells the Vta to connect the olfactory bulb (Ob) to the motor locomotive region (MLR), aiming the animal to the food scent.
The second shows orexin as a general food exploration signal. In contrast with the target-focused seeking, exploration is a random search.
The third is part of the eating circuit, where CGRP (an alarm neuropeptide) tonically inhibits eating, until AgRP (a hunger neuropeptide) disinhibits it [Essner et al. 2017].
Chiang, M. C., Bowen, A., Schier, L. A., Tupone, D., Uddin, O., & Heinricher, M. M. (2019). Parabrachial Complex: A Hub for Pain and Aversion. The Journal of neuroscience : the official journal of the Society for Neuroscience, 39(42), 8225-8230.
I’m considering exploring Braitenberg’s vehicles [Braitenberg 1984] for essay 14 in combination with the ideas from the archaoslug. The vehicles are a simple almost trivial design with surprisingly useful behavior. Each vehicle has a combination of sensor-motor pairs, taking advantage of the physical layout for the motor and sensor behavior.
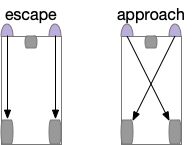
Here, the sensors detect light and directly drive the motor wheels. Vehicles with crossed signals approach the light, while vehicles with uncrossed signals avoid the light. Braitenberg also explores negative signals where the signals inhibit the motors, and additional signal-motor pairs for different senses. The value of the Braitenberg vehicles is showing how simple control circuits can form the basics of behavior.
Optic tectum as an example
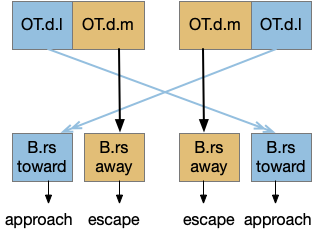
The optic tectum uses this dual-circuit architecture for approach and escape [Isa 2021]. The optic tectum is a midbrain optical and motor area responsible for much of vision in non-mammalian vertebrates and an understudied component of mammalian vision. In the OT, escape signals connect to ipsilateral (same side) motor neurons, and approach signals connect to a different set of motor neurons but crossing sides.
In the diagram, B.rs are reticulospinal motor neurons in the brainstem. OT.d.m is the medial optic tectum in the deep layers, and OT.d.l is the corresponding lateral. The OT shallow layers process optical information, and the deep layers drive motor actions. Stimulating the medial OT makes the animal escape and stimulating the lateral OT encourages approach. As a mnemonic, since approaching needs to aim at the target, its sensors need to be spread out (lateral), but escaping needs less precision and can rely on closer or merged (medial) sensors.
Because the Braitenberg architecture is so simple, I think it’s reasonable to imaging that primitive animals would quickly develop a similar pair of crossed and uncrossed systems as soon as neurons with specific connectivity were available, after the initial broadcast repeater nerved nets like in sea anemones (cnidaria). The dual systems mirrors the dual chemosensory and mechanosensory cell families in the archaoslug, which might have also encouraged split control.
Essay 14 pre-design
As a bilateral enhancement to the amoeboid archaoslug, I’m thinking of trying a ciliomotor slug with primitive obstacle avoidance but without any directed approach. Avoidance is a smaller evolutionary step because it can reuse the mechanosensory and nerve nets of the archaoslug, only adding a single crossed-pair of long-range neurons. After hitting an obstacle, muscle contractions turn the animal away from the obstacle.
The mucociliary sole remains the main locomotion and food detection. The slug still searches for food by grazing randomly on algae or bacterial mats, relying on browning motion to find food. There’s no tracking or approach system.
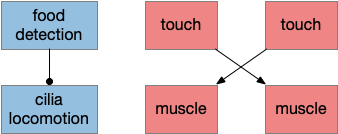
As mentioned above, the control systems for grazing locomotion and for obstacle avoidance are independent. Cilia locomotion is automatic with no neural control until the slug detects that it’s above food, when it stops. The locomotion direction is semi-random.
If the slug hits a wall, it contracts the side opposite the touch. This circuit is flipped from the Braitenberg vehicle, which has uncrossed signals for avoidance. The touch sensor activates the contralateral nerve net to contract the side muscle, and the slug turns toward the contracted side, away from the obstacle.
On motivation
Even in this trivial example, I think it’s useful to consider motivation in contrast with stimulus/response behavior. Since the basis of the word motivation is “to move,” it’s reasonable to use motivation as meaning moving force. So, motivation is a source of action without needing external stimulus. In the Braitenberg slug, the motivation is in the mucociliary sole itself, because it moves without external stimulus. If so, the motivation isn’t even neural; it’s just started by evolution.
The distinction of motivated vs non-motivated action is important in understanding the system. Knowing the sources of intrinsic motivation allows for tracing action from the source to its final result. As a design principle, adding self-motivation is more stable, because the animal is less likely to get stuck waiting for external stimuli to get started.
References
Braitenberg, V. (1984). Vehicles: Experiments in synthetic psychology. Cambridge, MA: MIT Press. “Vehicles – the MIT Press”
Isa, Tadashi, et al. “The tectum/superior colliculus as the vertebrate solution for spatial sensory integration and action.” Current Biology 31.11 (2021): R741-R762. https://doi.org/10.1016/j.cub.2021.04.001
[Arendt 2015] examines the fundamental divisions of the nervous system by looking at ancestral cell divisions in some of the earliest animals, specifically a multicellular amoeboid bottom-feeder that glides on a mucus foot like a slug. The archaoslug moves like an amoeba instead of a true slug because it’s body isn’t symmetrical bilateral: there’s no front or side. The Adrent study is interesting for the essays because it fundamentally splits chemosensory control (hypothalamic and olfactory) split from mechanosensory / optosensory sense and muscle brain at the cell type and developmental level.
In this proto-slug, cells have specialized into three major classes:
External skin with mechanosensory and optosensory cells
Internal digestive gut
Mucociliary sole with chemosensory and locomotion cells
The mucociliary sole moves with cilia gliding over mucus. Chemical sensors that detect food choose when to stop. A similar locomotive strategy is described in [Smith 2015] and [Senatore 2017] for the existing algae-grazing, disc-shaped animal Trichoplax, which lacks any nerves at all and only has six cells in total [Smith 2014].
Locomotion and food searching for the archaoslug is simple: stop when the chemosensors detects food (algae), and move in a random brownian direction when no food is available. A simple chemical sensor and a peptide-based broadcast system would suffice, as in Trichoplax. Because the bacterial mats may have dominated the Precambrian environment, the brownian motion pausing for food could work.
The ventral skin specialized into mechanosensory cells, optosensory cells, and contractile cells which developed into the first neurons and muscles. The animal can avoid obstacles and threats using nerve nets that broadcast and repeat signals, like the repeating nerve nets in cnidaria (jellyfish, sea anemone, and corals) [Seipel 2005]. Note, though, the control circuits for between locomotion (mucociliary sole) and muscles (skin/body contractions) are distinct, and don’t even coordinate. A sea anemone or a slug will contract when touched, but the sea anemone has no locomotion and the slug’s contraction isn’t its primary locomotion. Similarly, an archeoslug with primitive muscles might only use the muscles to avoid obstacles, contracting when it runs into something, but its primary motion remains the ciliary, non-muscular sole. Meaning, the locomotive drive (arrest, approach, avoid) is controlled independently from navigation (spatial obstacle avoidance.)
This division into three types influences all later cell development, because the initial decisions shape the later evolved cell types. Genetic signaling to choose between the three might have created a path dependent split between three types.
Discussion
This split between chemosensory sole and mechano- and opto-sensory skin and muscle obviously mirrors the vertebrate split between the limbic system (olfactory and hypothalamic) and optic tectum system, but with a different spin. The limbic system is generally described as a motivational and emotional center. The root word for both motivation and emotion is the Latin movere, to move, which has less baggage than either word. The mucociliary sole area does move and control movement, but it’s hardly an emotional center. But the mucociliary sole area isn’t unique in its control of motion, because the unrelated skin/muscle area controls navigation.
Treating the mind as independent, conflicting centers resembles Dawkins’ descriptions of genes in The Selfish Gene, where each gene works independently and in competition with others, and coordination only occurs for mutual benefit. The general idea of competing mental centers is also in Minsky’s Society of mind, and the idea is older than either. So, the value of the archaeoslug isn’t the general idea of a divided mind, but the specific division that occurred in evolution.
References
Arendt D, Benito-Gutierrez E, Brunet T, Marlow H. Gastric pouches and the mucociliary sole: setting the stage for nervous system evolution. Philos Trans R Soc Lond B Biol Sci. 2015 Dec 19;370(1684):20150286. doi: 10.1098/rstb.2015.0286. PMID: 26554050; PMCID: PMC4650134.
Dawkins, Richard. The Selfish Gene. Oxford University Press, 2006.
Minsky, Marvin. Society of mind. Simon and Schuster, 1988.
Senatore A, Reese TS, Smith CL. Neuropeptidergic integration of behavior in Trichoplax adhaerens, an animal without synapses. J Exp Biol. 2017 Sep 15;220(Pt 18):3381-3390. doi: 10.1242/jeb.162396. PMID: 28931721; PMCID: PMC5612019
Smith CL, Pivovarova N, Reese TS. Coordinated Feeding Behavior in Trichoplax, an Animal without Synapses. PLoS One. 2015 Sep 2;10(9):e0136098. doi: 10.1371/journal.pone.0136098. PMID: 26333190; PMCID: PMC4558020.
Smith CL, Varoqueaux F, Kittelmann M, Azzam RN, Cooper B, Winters CA, Eitel M, Fasshauer D, Reese TS. Novel cell types, neurosecretory cells, and body plan of the early-diverging metazoan Trichoplax adhaerens. Curr Biol. 2014 Jul 21;24(14):1565-1572. doi: 10.1016/j.cub.2014.05.046. Epub 2014 Jun 19. PMID: 24954051; PMCID: PMC4128346.