The ascidian circuit in essay 30 had an interesting dopamine subcircuit that looks like an indirect search, where the ascidian coronet cells modulate the underlying phototaxis and geotaxis circuits. While the function of the coronet cells is unknown, if these cells are another seeking system like following an odor, then the coronet sub circuit follows odor by modulating different seek circuits: phototaxis and geotaxis.
Ascidian analogy
Tunicates are the closest non-vertebrate chordates evolutionarily, but they have developed in vastly different directions from the vertebrates, and likely very differently from the shared common ancestor [Holland 2015]. The ascidian tunicates, which are the most studied tunicates, live their asul life as sessile filter feeders like sponges. Their eggs hatch in only 20 hours and their brief tadpole form lasts only for a few hours, just enough to swim and disperse to find a likely permanent settlement place. Their locomotive strategy is to swim up using geotaxis in the morning and swim down using phototaxis in the afternoon. If they’re lucky enough to find a ledge, they swim up into the ledge’s shadow to settle because hanging like a bat from a ledge offers more protection from some predators than resting on the ocean floor [Zega et al 2006].
As would be expected from a 20-hour brain, the navigation circuit is fairly simple. There are two distinct action paths, one for geotaxis using a heavy pigment cell and one for phototaxis using photoreceptors and another pigment cell as a shadow to provide photo-directionality. The two action paths are connected, where dimming produces upward swimming [Bostwick et al 2020].
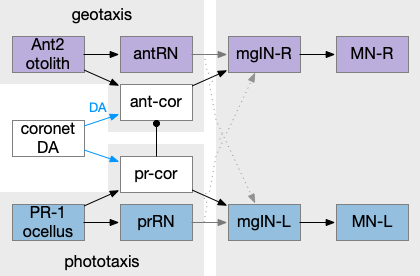
Ascidian tadpole sub circuit for geotaxis and phototaxis. The horizontal neurons are the main action paths. The coronet DA cells modulate the action paths.
In the above diagram, the geotaxis action path starts from the otolith (“ear stone”) receptor ant2, which is functionally similar to the vestibular system (but not related), passes input to antenna relay neurons (antRN) and then to the right side motor neurons (mgIN-R and MN-r) [Ryan et al 2016]. Similarly, the phototaxis action path starts from the ocellus (eyespot) to the phototaxis relay (prRN) and to the left motor neurons, providing an opposing direction from geotaxis. Importantly for the following discussion, each path has a weak connection to the opposite direction, possibly to add some stochasticity to the movement to improve dispersion of the many tadpoles.
The function of the coronet cells is unknown, although they have some genetic connection the palp sensory cells [Cao et al 2019]. Other papers compare the corona cells to dopamine cells in the hypothalamus and Ob (olfactory bulb) [Horie et al 2018] or ancestral photo-hypothalamus and retina [Sharma et al 2019], possibly related to the fish saccus vasculosus area of the hypothalamus, responsible for some circadian behavior. However, the ascidian tadpole has lost circadian clock genes, which argues against circadian timing [Chung et al 2023]. The coronet cells can accumulate serotonin and the DA might promote onset of metamorphosis [Razy-Kraika et al 2012]. So, the coronet may be involved in triggering metamorphic changes at twilight, which causes the tadpole to dive to deeper waters [Lemaire et al 2021].
Whatever the source, the interesting thing about the circuit is that it’s an indirect modulation of underlying taxis action paths. The action of the coronet is gating or modulatory. While this coronet circuit is not homologous to the basal ganglia, using it as an analogy may be useful. For example, dopamine is a sleep / wake signal for the basal ganglia [Vetrivelan et al 2010]. Because low dopamine reduces basal ganglia activity both at the striatum input layer and the Snr (substantia nigra pars reticulata) output layer, it’s an effective sleep controller.
Indirect chemotaxis
Consider indirect chemotaxis, where the animal is seeking toward the odor, but the underlying action path is phototaxis or geotaxis, like the ascidian circuit above. If the animal detects an odor, it increases the current direction. In other words, the current direction is toward or near a food odor. This strategy is like the e. coli tumble-and-run strategy, where the bacteria runs further when the odor gradient is increasing.
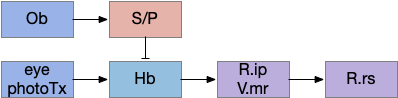
Consider the basal ganglia as an analogy. For example, Ob has some dopamine interneurons (Ob.sac – short axis cells) that project to S.ot (olfactory tubercle) [Burton 2017], a portion of the stratum focused on olfactory input. For the corollary of the phototaxis path, consider the Hb.m (medial habenula) phototaxis path [Zhang et al 2017].
Hypothetical indirect seek circuit where chemotaxis uses an underlying phototaxis to hunt for food. Hb (habenula), Ob (olfactory bulb), P (pallidum), R.ip (interpeduncular nucleus), R.rs (reticulospinal motor neurons), S (striatum), V.mr (median raphe).
When the odor is detected, Ob enables the basal ganglia, which enhances the phototaxis path. If the odor isn’t detected, the default semi-suppressed behavior means the direction is semi-random. This indirect control would allow for seeking odor when the underlying navigation is phototaxis and geotaxis.
Discussion
After writing this description. I think this model may be a bit sketch for something like chemotaxis, although it’s a reasonable model for sleep. Because I’m not sure the idea is likely to be productive, I’m holding off on doing any implementation, but writing down the description in case it makes sense later.
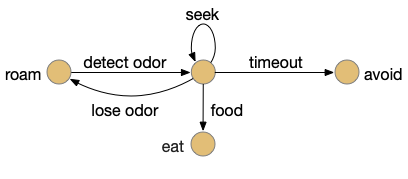
Let’s return to the task of essay 16 on give-up time in foraging, which covered food search with a timeout. At first the animal uses a general roaming search and if it smells a food odor, it switches to a targeted seek following the odor with chemotaxis. If the animal finds food in the odor plume, it eats the food, but if it doesn’t find food, it will eventually give up and avoid the local area before returning to the roaming search.
Search state machine. Roam is the starting state, switching to seek when it detects odor, and switching to avoid after a timeout.
For another attempt at the problem, let’s take the striatum (basal ganglia) as implementing the timeout portion of this task using the neurotransmitter adenosine as a timeout signal and incorporating the multiple action path discussion from essay 30 on RTPA. Adenosine is a byproduct of ATP breakdown and is a measure of cellular activity. With sufficiently high adenosine, the striatum switches from the active seek path to an avoidance path. These circuits are where caffeine works to suppress the adenosine timeout, allowing for longer concentration.
Mollusk navigation
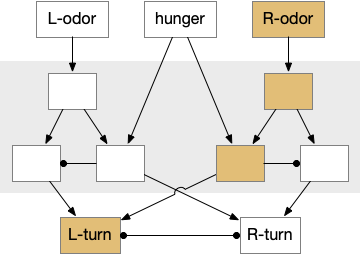
As mentioned in essay 30, the mollusk sea slug has a food search circuit with a similar logic to what we need here. The animal seeks food odors when it’s hungry, but it avoids food odors when it’s not hungry [Gillette and Brown 2015].
Mollusk food search circuit, illustrating a hunger-modulated switchboard. When the animal is not hungry, the switchboard reverses the odor to motor links turning it away from food.
This essay uses the same idea but replaces the hunger modulation with a timeout. When the timeout occurs, the circuit switches from a food seek action path to a food avoid action path.
Odor action paths
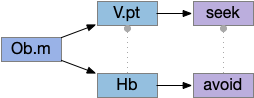
Two odor-following actions paths exist in the lamprey, one using Hb.m (medial habenula) and one using V.pt (posterior tuberculum). The Hb.m path is a chemotaxis path following a temporal gradient. The V.pt path projects to MLR (midbrain locomotor region), but The lamprey Ob.m (medial olfactory bulb) projects to both Hb.m (medial habenula) and to V.pt (posterior tuberculum), which each project to different locomotor paths [Derjean et all 2010], Hb.m to R.ip (interpeduncular nucleus) and V.pt to MLR (midbrain locomotor region). The zebrafish also has Ob projections to Hb and V.pt [Imamura et al 2020], [Kermen et al 2013].
Dual odor-seeking action paths in the lamprey and zebrafish. Hb (habenula), Ob.m (medial olfactory bulb), V.pt (posterior tectum).
Further complicating the paths, the Hb.m itself contains both an odor seeking path and an odor avoiding path [Beretta et al 2012], [Chen et al 2019]. Similarly Hb.m has dual action paths for social winning and losing [Okamoto et al 2021]. So, this essay could use the dual paths in Ob.m instead of contrasting Ob.m with V.pt, but the larger contract should make the simulation easier to follow.
This essay’s simulation makes some important simplifications. The Hb to R.ip path is a temporal gradient path used for chemotaxis, phototaxis and thermotaxis. In a real-world marine environment, odor diffusion and water turbulence is much more complicated, producing more clumps and making a simple gradient ascent more difficult [Hengenius et al 2012]. Because this essay is only focused on the switchboard effect, this simplification should be fine.
Striatum action paths with adenosine timeout
The timeout circuit uses the striatum, which has two paths: one selecting the main action, and the second either stopping the action, or selecting an opposing action [Zhai et al 2023]. The two paths are distinguished by their responsiveness to dopamine with S.d1 (striatal projection with D1 G-s stimulating) or S.d2 (striatal projection with D2 G-i inhibiting) marking the active and alternate paths respectively. This model is a simplification of the mammalian striatum where the two paths interact in a more complicated fashion [Cui et al 2013].
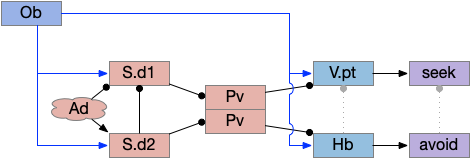
Essay odor seek with timeout circuit. The seek path flows from Ob, through S.d1 to P.v to V.pt. The avoid path flows from Obj, though S.d2 to Pv. to Hb. Ad (adenosine), Hb (habenula), Ob (olfactory bulb), Pv (ventral pallidum), S.d1 (striatum D1 projection neuron), S.d2 (striatum D2 projection neuron), V.pt (posterior tuberculum)
As mentioned, the two actions paths are the seek path from Ob to V.pt and the avoid path from Ob to Hb. For the timeout and switchboard, the Ob has a secondary projection to the striatum. Although this circuit is meant as a proto-vertebrate simplification, Ob does project to S.ot (olfactory tubercle) and to the equivalent in zebrafish [Kermen et al 2013].
The timeout is managed by adenosine, which is a neurotransmitter derived from ATP and a measure of neural activity. The striatum has three sub-circuits for this kind of functionality, which I’ll cover in order of complexity.
S.d1 and adenosine inhibition
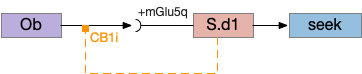
The first circuit only uses the direct S.d1 path and adenosine as a timeout mechanism. When the animal follows an odor, the Ob to S.d1 signal enables the seek action. As a timeout, ATP from neural activity degrades to adenosine and the buildup of adenosine is a decent measure of activity over time. The longer the animal seeks, the more adenosine builds up. Of the Ob projection axis contains an A1i (adenosine G-i inhibitory) receptor, the adenosine will inhibit the release of glutamate from Ob, which will eventually self-disable the seek action.
S.d1 action path inhibited by adenosine buildup as a timeout. A1i (adenosine G-i inhibitory receptor), Ad (adenosine), mGlu5q (metabotropic glutamate G-q receptor), Ob (olfactory bulb), S.d1 (D1-type striatal projection neuron)
In practice, the striatum uses astrocytes to manage the glutamate release. An astrocyte that envelops the synapse measures glutamate release with an mGlu5q (metabotropic glutamate with G-q/11 binding) receptor and accumulates internal calcium [Cavaccini et al 2020]. The astrocyte’s calcium triggers an adenosine release as a gliotransmitter, making the adenosine level a timeout measure of glutamate activity. The presynaptic A1i receptor then inhibits the Ob signal. The timeframe is on the order of 5 to 20 minutes with a recovery of about 60 minutes, although the precise timing is probably variable. Interestingly, the time-out is a log function instead of linear measure of activity [Ma et al 2022].
This circuit doesn’t depend on the postsynaptic S.d1 firing [Cavaccini et al 2020], which contrasts with the next LTD (long term depression) circuit which only inhibits the axon if the S.d1 projection neuron fires.
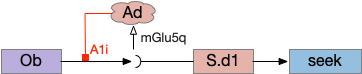
S.d1 presynaptic LTD using eCB
S.d1 self-activating LTD uses retrotransmission to inhibit its own input using eCB (endocannabiniods) as a neurotransmitter. Like the astrocyte in the previous circuit, S.d1 uses a mGlu5q receptor to trigger eCB release, but also require that S.d1 fire, as triggered by NMDA glutamate receptor. The axon receives the eCB retrotransmission with a CB1i (cannabinoid G-i inhibitory) receptor and trigger presynaptic LTD [Shen et al 2008], [Wu et al 2015]. Like the previous circuit, the timeframe seems to be on the order of 10 minutes, lasting for 30 to 60 minutes.
S.d1 LTD circuit. A coincidence of glutamate detection with mGlu5q and S.d1 activation with NMDA triggers eCB release, which activates CB1i leading to presynaptic LTD. CB1i (cannabinoid G-i inhibitory receptor), mGlu5q (glutamate G-q receptor), Ob (olfactory bulb), S.d1 (striatum D1-type projection neuron).
This circuit inhibits itself over time without using adenosine or astrocytes. In the full striatum circuit, high dopamine levels suppress this LTD suppression, meaning that dopamine inhibits the timeout [Shen et al 2008].
The next circuit adds the S.d2 path, which uses adenosine and self-activity to trigger postsynaptic LTD.
S.d2 postsynaptic LTP via A2a.s
Consider a third circuit that has the benefits of both previous circuits because it uses adenosine as a timer managed by astrocytes and is also specific to postsynaptic activity. In addition, it allows for a second action path, changing the circuit from a Go/NoGo system to a Go/Avoid action pair. This circuit uses LTP (long term potentiation) on the S.d2 striatum neurons.
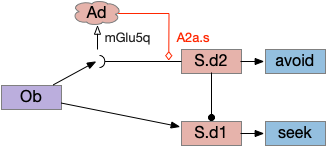
Timeout circuit using postsynaptic LTD at the S.d2 neuron and adenosine as a timeout signal. As adenosine accumulates, it stimulates S.d2, which both disables S.d1 and drives the avoid path. A2a.s (adenosine G-s stimulatory receptor), Ad (adenosine), mGlu5q (glutamate G-q metabotropic receptor), Ob (olfactory bulb), S.d1 (striatum D1-type projection neuron), S.d2 (striatum D2-type projection neuron)
When the odor first arrives, Ob activates the S.d1 path, seeking toward the odor. S.d1 is activated instead of S.d2 because of dopamine. In this simple model, the Ob itself could provide the initial dopamine like c. elegans odor-detecting neurons or the tunicate’s coronal cells or the dual glutamate and dopamine neurons in Vta (ventral tegmental area).
As time goes on, adenosine from the astrocyte builds up, which activates the S.d2 A2s.a (adenosine G-s stimulatory receptor) until it overcomes dopamine suppression and increases the S.d2 activity with LTP [Shen et al 2008]. Once S.d2 activates, it suppresses S.d1 [Chen et al 2023] and drives the avoid path.
The combination of these circuits looks like it’s precisely what the essay needs.
Simulation
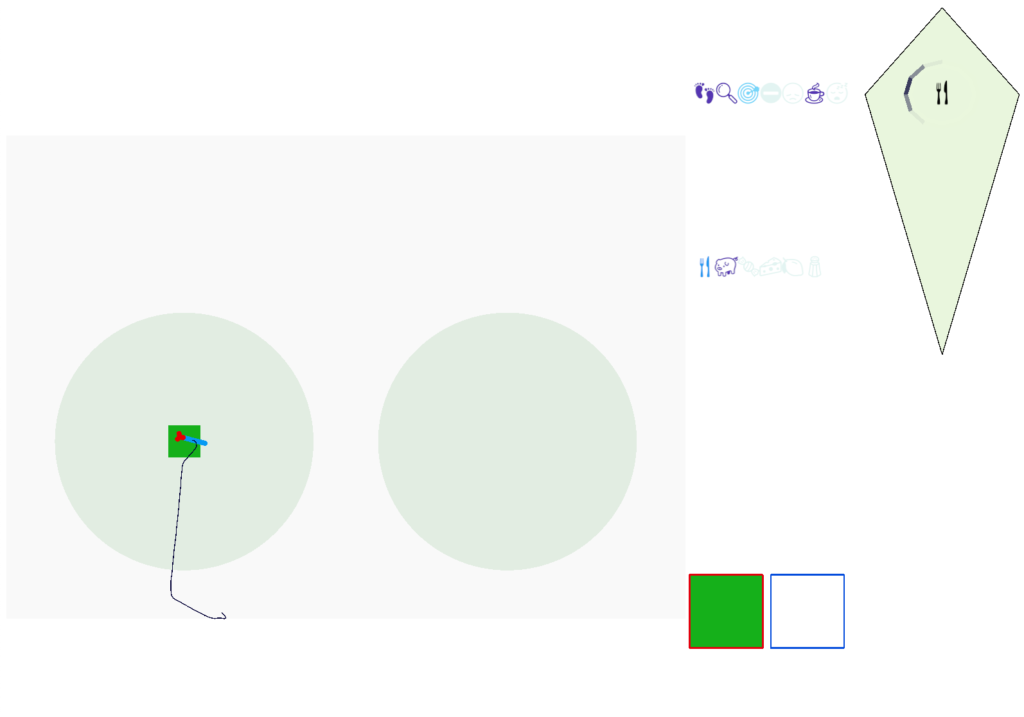
In the simulation, when the animal is hunting food and finds a food odor plume, it directly seeks toward the center and eats if it find food. In the screenshot below, the animal is eating.
Simulation showing the animal eating food after seeking the odor plume.
Satiation disables the food seek. This might sound obvious, but hunger gating of food seeking requires specific satiety circuits to any seek path that’s food specific, which means the involvement of H.l (lateral hypothalamus) and related areas like H.arc (arcuate hypothalamus) and H.pv (periventricular hypothalamus). And, of course, the simulation requires simulation code to only enable food odor seek when the animal is searching for food.
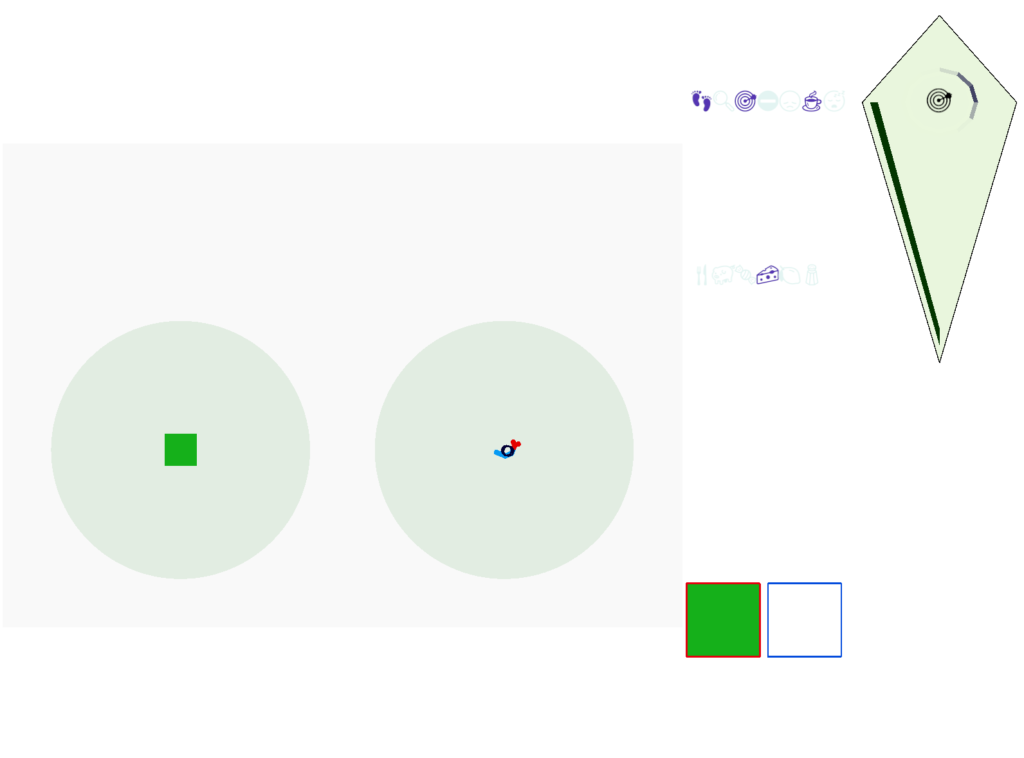
The next screenshot shows the central problem of the essay, when the animal seeks a food odor but there’s no food at the center.
Screenshot showing the animal stuck in the middle of the food odor plume before the timeout.
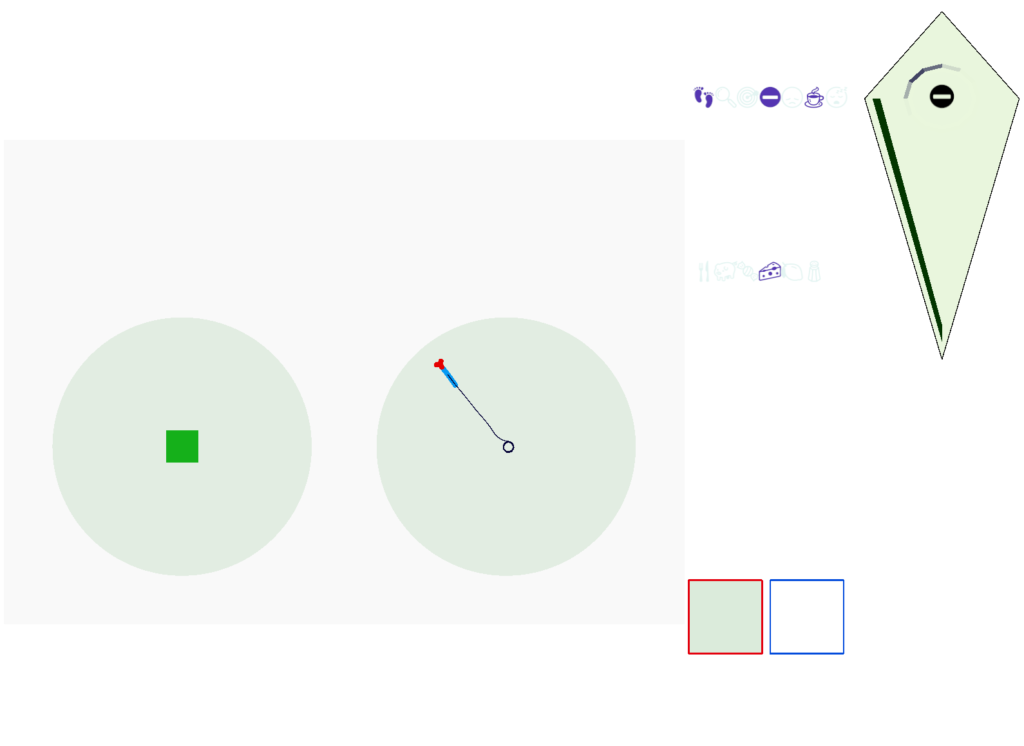
Without a timeout, the animal circles the center of the food odor plume endlessly. After a timeout, the animal actively leaves the plume and avoid that specific odor until the timeout decays.
Screenshot showing the animal escaping from the odor plume after the timeout.
This system is somewhat complex because of the need for hysteresis. A too-simple solution with a single threshold can oscillate, because as soon as the animal starts leaving the timeout decays, which then re-enables the food-seek, which then quickly times out, repeating. Instead, the system needs to make re-enabling of the food seek more difficult after a timeout.
But that adds a secondary issue because if food seek is a lower threshold, then the sustain of seek needs to raise the threshold while the seek occurs. So, the sustain of seek needs a lower threshold than starting seek. This hysteresis and seek sustain presumably needs to be handled by the actual striatum circuit.
Discussion
I think this essay shows that using the stratum for an action timeout for food seek is a plausible application. The circuit is relatively simple and is effective, improving search by avoiding failed areas.
However, the simulation does raise some issues, particularly hysteresis problem. If the striatum does provide a timeout along these lines, it must somehow solve the hysteresis problem. While the animal is seeking, the ongoing LTP/LTD inhibition should use a high threshold to stop seeking, but once avoidance starts, there needs to be a high threshold to return to seeking to avoid oscillations between the two action paths.
Because LTD/LTP is a relatively long chemical process (minutes) internal to the neurons, as opposed to an instant switch in the simulation, the delay itself might be sufficient to solve the oscillation problem. It’s also possible that some of the more complicated parts of the circuit, such as P.ge (globus pallidus) and its feedback to the striatum or H.stn (subthalamic nucleus) might affect the sustain of seek or breaking it and so control the hysteresis problem.
The simulation also reinforced the absolute requirement that action paths need to be modulated by internal state like hunger. For the seek paths, both Hb.m and V.pt are heavily modulated by H.l and other hypothalamic hunger and satiety signals.
As expected, the simulation also illustrated the need for context information separate from the target odor. While the food odor is timed out, the animal can’t search the other odor plume because this essay’s animal can’t distinguish between the odor plumes, and therefore avoids both odors. With a long timeout and many odor plumes, this delays the food search. A future enhancement is to add context to the timeout. If the animal can timeout a specific odor plume, it can search alternatives even if the food odor itself is identical.
Unsurprisingly since essay 26 was a first cut at selective attention, it exposed a number of problems with both the neuroscience and the simulation model itself.
Specific give up
The current give up circuit is a global circuit, which doesn’t depend on the current stimulus. For this essay, the animal has two potential and because the give up is global, when the animal gives up, it gives up on both odors.
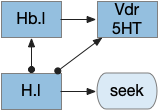
Global give-up circuit for olfactory seek. H.l lateral hypothalamus, Hb.l lateral habenula, Vdr dorsal raphe, 5HT serotonin.
An improvement would be a cue-specific give up capability. When the animal gives up on odor A, it should investigate odor B. Instead it gives up on both. I need to add some mechanism to create a cue-specific give up capability.
As a possible neural analog, the adenosine receptor can work as a local give-up circuit by integrating neural activity. Since adenosine is essentially a waste produce from neural activity, long activity will accumulate adenosine. The A1 adenosine receptor detects the adenosine and inhibits activity, since it’s a Gi receptor.
Olfactory complexity and attention
The essay’s odor model is extremely oversimplified, because odor receptors are feature detectors, not molecule receptors, and odors are combinations of molecules. Since a specific odor is a combination of features, P.bf (basal forebrain) can’t be a simple winner-take-all inhibitory circuit as implemented in this essay. Instead, attention needs to be a set of features that excludes the distractor odor’s features.
Olfactory gamma and beta
Although the essay treats the olfactory bulb data as direct signals, oscillations are a major feature of the olfactory bulb. Strong odors trigger gamma (40-100Hz) signals in Omt (mitral/tufted output cells), enhanced by ACh (acetylcholine) from P.bf. Feedback from O.pir (olfactory piriform cortex) triggers beta (15-30Hz) oscillations. In addition, interactions with breathing in mammals synchronized with theta (4-10Hz). Although, in the last case, since the simulation animal is aquatic, breathing isn’t an appropriate synchronizer.
Temporal gradient seek issues
Odor seeking in essay 26 uses temporal gradient descent modulated by head direction in Hb.m (medial habenula) and B.ip (interpeduncular nucleus). The animal combines its head direction with the temporal gradient to estimate the odor direction, and it saves the result as a goal vector. As the animal turns, it can improve the direct estimate. In the phototaxis example of essay 25, the saved goal vector direction helped with intermittent data, where it could remember the light location for a few seconds.
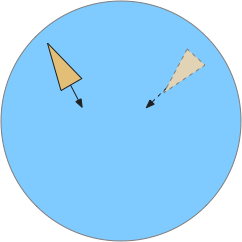
Problems with the current odor direction. A quick switch in location incorporates data from the old direction, leading to an incorrect estimate.
However, the system as implemented in the model is extremely limited. It can’t truly triangulate to locate the odor, but can only improve the single direction. In the diagram above, the animal can only select one of the two vectors as an estimate. It can’t combine the two into a better estimate of the center. Also, in the diagram, the earlier estimate is no longer useful because the animal has moved.
Now, the issue might be purely in the simulation. If B.ip and Vdr (dorsal raphe serotonin) are calculating this kind of estimate, it’s likely their computation is better than the current simulation.
The selection is a trade off where a stronger gradient is likely a better estimate, but if the animal moves too far from the earlier sample, the old direction is no longer relevant. Since the animal lacks the sophistication of an allocentric map to resolve the discrepancy, it discards the old value.
The current implementation decays the old estimate to allow newer estimates to overwrite it even if the later gradient is weaker. Essentially the memory is like a leaky integrator, as is appropriate for placing it in the serotonin neurons and/or associate glia with short term (5s) memory as in simple zebrafish motor memory [Dragomir et al 2020].
Bayesian updates
In a future essay, it might be interesting to explore this issue to see if a simple Bayesian system could be implemented in low-complexity circuits, where stronger data would update the current model more than the current model.
Self motion and gradient vectors
When the animal is turning, the running average no longer represents a straight line. For the gradient vector, the system assumes the recent average was measured along the current head direction, but turns violate this assumption. To avoid miscalculating gradient vectors, the animal should suppress measurement during turns.
Swimming and theta
The gradient seek issues above are compounded with swimming with a fixed head. Early vertebrates would have had a fixed head like sharks, meaning that each swimming stroke would move the head from side to side. That sideways movement would affect the odor gradient and head direction.
Inconsistent head vs body direction and odor measurement while swimming with a fixed head.
A simplistic fix would take an odor gradient sample only on each swim stroke, only reporting at the stroke end for consistency and to average from the beginning of the stroke to the end. That solution would give a consistent measurement in a reasonably consistent direction, as opposed to sampling randomly in a cycle.
Log encoding vs linear encoding
For simplicity, I’e used linear encoding for signals in the essays, because the basic functional architecture remains the same, and the simulation isn’t precise enough to need more complexity. But for odors, the dynamic range between a single molecule detection and an overpowering odor doesn’t scale well with a linear representation.
In particular, the odor weight from the simple distance gradient, together with above mentioned temporal gradient issues might be better modeled with a log signal. Basically, the issue I raised above with gradient vector sampling might be more tractable with a different encoding, and log encoding might make the actual neural circuit less finicky than the current linear model.
Seek mode switching
The essay’s simulation lacks a specific mode switching circuit. In vertebrates the peptide core (hypothalamus, PAG, B.pb area) switches action modes from roaming to seek to eating to rest and sleep. These modes are motivated and depend on internal needs and scheduling impulses programmed by evolution.